![]()
5 février 2009 - Journée d'étude ASFC « Vérone
- Ombres & Lumières »
Génétique
et Sélection
au 9ème Congrès Mondial de Cuniculture
par
Hervé GARREAU* et Jacques HURTAUD**
* INRA, SAGA, Centre de recherches de Toulouse, 31326 Castanet-Tolosan
Cedex
** Grimaud Frères Sélection, La Corbière 49450
Roussay
Introduction
Quarante-cinq communications (un rapport de synthèse et 44 communications courtes) ont été présentées lors de la session " Génétique", soit vingt-trois de plus qu'au dernier congrès mondial. L'Espagne est de loin la mieux représentée avec 14 communications contre 6 pour la France, 5 pour l'Italie, 3 pour la Hongrie. L'ensemble de ces contributions peut s'articuler autour de 3 volets : variabilité génétique et sélection des caractères, génétique moléculaire, caractérisation de populations locales.
Les méthodes classiques de la génétique quantitative (modèle polygénique et estimation des effets génétiques basés sur la régression des performances et l'information des apparentements) restent les outils privilégiés des généticiens lapins. Avec 19 communications, ce domaine d'étude est le plus représenté de la session.
1. Sélection pour l'homogénéité des performances

H. Garreau et J. Hurtaut lors de leur présentation
|
Très innovante dans le domaine de la sélection animale, cette méthode donne des résultats très prometteurs et a fait l'objet de 4 communications. Elle suppose que les caractères sont contrôlés par deux groupes de gènes, un groupe contrôlant la moyenne du caractère (ex : poids des lapereaux à la naissance) et l'autre sa variabilité (ex : variabilité de ces poids intra-portée) et elle estime pour chaque individu deux valeurs génétiques distinctes. C'est en sélectionnant les animaux sur la valeur génétique pour la variabilité du caractère qu'il est possible d'homogénéiser un caractère. Bolet et al. ont présenté les résultats de 7 générations de sélection divergente pour la variabilité des poids de naissance du lapereau, pour une expérience dont les premiers résultats avaient été donnés par Garreau et al. au précédent congrès mondial : en comparaison avec la lignée hétérogène, la lignée homogène présente une variabilité des poids à la naissance plus faible, une mortinatalité et une mortalité naissance sevrage plus faible, une prolificité à la naissance et au sevrage plus élevée. Les femelles de la lignée homogène ont également des cornes utérines significativement plus longues et plus à même de s'allonger, offrant vraisemblablement plus d'espace pour l'accueil des fœtus. Il n'existe malheureusement pas de lignée témoin pour vérifier la symétrie de la réponse. En revanche des congélations d'embryons ont été réalisées dans ce but chez le sélectionneur Grimaud Frères Sélection avant de mettre en place cette méthode de sélection dans la lignée AGP22. Ces embryons permettront de constituer une population témoin pour vérifier la réponse à la sélection après 5 années de sélection (en 2011). |
 |
Emboîtant le pas de l'INRA les généticiens de l'Université de Valence (Espagne) ont débuté une expérience de sélection sur la variabilité de la taille de portée à la naissance. Ibáñez-Escriche et al. ont présenté les paramètres génétiques des 2 composantes génétiques du caractère : les variances génétiques de la moyenne et de la variabilité sont respectivement de 0,77 et 0,12 avec une corrélation favorable (-0,74) entre les 2 composantes. Les résultats préliminaires obtenus après une génération de sélection divergentes pour la variance de la taille de portée confirment ces paramètres (Argente et al., Espagne) : La variance de la lignée haute est significativement plus élevée que celle de la lignée basse. La diminution de la variance dans la lignée baute s'accompagne d'une augmentation de la moyenne de la taille de portée (+0,4 lapereau nés totaux). Si ces résultats se confirment, les généticiens auront à disposition une méthode de sélection très intéressante pour réduire la variabilité de la taille de portée tout en améliorant son niveau moyen. Dans le même domaine, Ibáñez-Escriche et al. ont estimé les paramètres génétiques des 2 composantes génétiques de la capacité utérine à partir des mesures obtenues pendant 10 générations de sélection : les variances génétiques de la moyenne et de la variabilité sont respectivement de 0,82 et 0,16 avec, encore, une corrélation favorable (-0,74) entre les 2 composantes.
Deux équipes espagnoles ont produit chacune 2 articles pour cette famille de caractères. Dans un premier article (I) Lavarra et al. donnent les valeurs d'héritabilité suivantes : volume d'éjaculat 0,091, concentration 0,053, nombre total de spermatozoïdes (spz) par éjaculat 0,097, pourcentage de spz avec un acrosome intègre 0,247, pourcentage de spz avec anomalie morphologique 0,643, longueur de la tête 0,309, largeur de la tête 0,341, surface de la tête 0,279, périmètre de la tête 0,105 , ellipticité de la tête 0,508, rugosité de la tête 0,518, élongation de la tête 0,220 et régularité de la tête 0,067. Les corrélations génétiques de ces caractères avec la vitesse de croissance en engraissement sont négatives pour la concentration (-0,69), le nombre total de spz (-0,36), la largeur de la tête (-0,617), la surface de la tête (-0,617) indiquant que la sélection pour la vitesse de croissance peut dégrader le niveau génétique de ces caractères. Dans un second article (II) les mêmes auteurs donnent les valeurs d'héritabilité des caractères de motilité de la semence : pourcentage de spz motiles 0,16, vélocité moyenne 0,19, index de linéarité 0,12, amplitude de déplacement latéral 0,07. Les corrélations génétiques de ces caractères de motilité avec la vitesse de croissance en engraissement sont également défavorables, modérées pour la plupart excepté pour le pourcentage de spermatozoïdes motiles (-0,93). En résumé les héritabilités moyennes à fortes des caractéristiques de la semence laissent envisager la possibilité d'une amélioration de ces caractères par la sélection. La sélection simple pour la croissance en engraissement tend à dégrader la plupart de ces caractéristiques.
Piles et al. ont décrit une étude visant à estimer les effets génétiques de la semence du mâle sur la fertilité (F) (article 1) et sur la taille de la portée (TP) (article 2) pour 2 niveaux de concentration de la semence inséminée (10 x 106 spz/ml et 40 x 106 spz/ml). L'objectif était de vérifier si la variabilité individuelle des mâles était différente pour des niveaux de dosage de la semence différents. Les paramètres génétiques des caractères F10, F40, TP10 et TP40 ont été estimés, en considérant les caractères comme différents pour les 2 niveaux de dosage et en estimant l'interaction génotype x dosage. L'effet du dosage était significatif, à la fois pour la fertilité (0,46 vs 0,59) et pour la prolificité (7,2 vs 8,5). Cependant les héritabilités de F10 et de F40 (0,08 et 0,09), d'une part et de TP10 et TP40 (0,08 et 0,06) d'autre part n'étaient pas significativement différentes et la valeur de l'interaction génotype x environnement était négligeable pour les 2 caractères. Ces résultats démontrent que les gènes qui agissent sur F10 (réciproquement TP10) sont les mêmes que ceux qui agissent sur F40 (réciproquement TP40). La sélection pour améliorer la composante génétique mâle de la fertilité et de la prolificité peut donc être réalisée indépendamment du niveau de dosage de la semence dans l'intervalle étudié.
L'équipe de l'université de Kaposvar (Hongrie) a de nouveau présenté des études basées sur l'utilisation de tomographe (rayons X) dans la lignée Pannon blanche. Szendrö et al. ont décrit une expérience de sélection divergente pour le volume de la cuisse mesuré par tomographie. La lignée P était sélectionnée pour augmenter le volume de la cuisse et la lignée M pour le réduire. Après 2 générations de sélection des différences significatives ont été observées entre les 2 lignées : la quantité d'aliment ingérée (128 g/j vs 138 g/j) et l'indice de consommation (2,81 vs 3,01) étaient meilleurs dans la lignée P que dans la lignée M ; la proportion de tractus gastro intestinal (16,7 % vs 18,1 %) et la proportion de gras perirénal (1, 90 vs 2,40) et periscaplulaire (0,49 vs 1,07) étaient plus faibles dans la lignée P tandis que la proportion de partie arrière de la carcasse (38,2 vs 36, 3) étaient plus élevée dans la lignée P. Ces résultats indiquent que la sélection pour le volume de la cuisse améliore non seulement les caractères de carcasse mais également la quantité d'aliment ingérée et l'indice de consommation. Selon Gyovai et al., le noyau de sélection de la lignée Pannon est sélectionné en 2 étapes : 30 à 40 % des candidats à la sélection sont choisis sur leur valeur génétique de croissance (GMQ) et l'épaisseur de leur cuisse est mesurée par tomographie. La sélection finale combine les valeurs génétiques des 2 caractères Les héritabilités du GMQ et du volume de la cuisse (EC) sont respectivement égales à 0,27 et à 0,21. La corrélation génétique entre les 2 caractères est faiblement favorable (0,14). Le progrès génétique estimé par le BLUP était de 1 g par année pour le GMQ (2,4 % de la moyenne du caractère) et de 4 cm3 par année pour l'épaisseur de la cuisse (1,2 % de la moyenne du caractère).
Pascual et al. (Espagne) ont étudiés les effets de la sélection pour la vitesse de croissance sur la croissance relative des organes, des tissus et des morceaux de découpe dans la lignée espagnole R. La liaison de ces caractères avec le poids a été analysée par le coefficient d'allométrie de Huxley (k) tandis que leur degré de maturité a été analysé par le coefficient d'allométrie de Butterfield (q). Les valeurs obtenues pour ces coefficients ont permis de repréciser le degré de maturité relatif de chacun de ces caractères mais aucun effet de la sélection sur ces coefficients n'a été mis en évidence pour l'ensemble des caractères étudiés.
Les résultats
d'une expérience de sélection divergente pour la longévité
de la lapine ont été présentés par Garreau
et al. (France). La sélection a porté sur le nombre d'inséminations
artificielles en utilisant la méthode de l'analyse de survie. Après
une génération de sélection, une différence
significative de longévité (+0,92 IA soit 39 jours) a été
mesurée entre les deux lignées. Hormis pour le nombre de
nés totaux plus élevé dans la lignée basse,
Il n'y avait pas de différence entre les deux lignées pour
les caractères de reproduction enregistrés à chaque
mise-bas. Cette expérience a démontré la faisabilité
d'une amélioration génétique pour améliorer
la durée de la carrière reproductive et la survie des femelles.
La méthode est aujourd'hui appliquée par le sélectionneur
Grimaud Frères Sélection.
Pascual et al. ont comparé l'efficacité alimentaire des
lapines multipares en lactation (période J13 à J16) de la
lignée V, sélectionnée pour la taille de portée
au sevrage, et de la lignée " longève " LP sélectionnée
pour la longévité. Les femelles de la lignée LP ont
consommé 22 g de matière sèche par jour en plus par
rapport à la lignée V. Le coefficient de digestibilité
était plus faible dans la lignée LP pour la matière
sèche, la matière organique et l'énergie brute (2,3,
2,5 et 2,1 % respectivement). Ces résultats montrent que la sélection
pour la taille de portée au sevrage a contribué à
sélectionner des femelles plus efficaces pour l'utilisation des
ressources nutritionnelles que la sélection pour la longévité.
5. Résistance aux troubles digestifs
|
Le thème de la résistance génétique aux maladies n'a fait l'objet que d'une seule communication, en dépit de son intérêt pour la filière. Garreau et al. ont élaboré un indice binaire (0/1), à partir des observations en routine de signes d'entéropathie (absence/présence) de l'ensemble des animaux contrôlés en engraissement depuis 1998 de la lignée AGP39 du sélectionneur Grimaud Frères Sélection. Les auteurs ont estimé les paramètres génétiques de cet indice et des caractères couramment sélectionnés dans cette lignée : poids à 63 jours, rendement de carcasse, taux de gras périrénal. Les héritabilités des caractères étaient respectivement de 0,08 , 0,36 , 0,24 et 0,64. Les corrélations génétiques entre l'indice de résistance, d'une part, et le poids à 63 jours et le rendement de carcasse, d'autre part, étaient négatives et donc favorables (-0,19 et -0,34 respectivement). La corrélation génétique entre l'indice de résistance et le taux de gras périrénal était négligeable (-0,07). Ces paramètres démontrent qu'il est possible de sélectionner les animaux pour la résistance aux troubles digestifs non spécifiques, à partir d'enregistrements sanitaires en routine, sans compromettre l'amélioration des caractères de production. Des protocoles de sélections divergentes sont à l'étude pour mieux comprendre les mécanismes de résistance. Ce nouveau critère a d'ores et déjà été introduit dans l'objectif de sélection dans une lignée commerciale du sélectionneur.
|
En bleu :
héritabilité des caractères et en
noir : correlations génétiques entre caractères.
|
Laborda et al. (Espagne)
ont décrit les résultats de 6 générations
de sélection pour le taux d'ovulation dans une population expérimentale.
Les lapines (80 par génération) étaient endoscopées
pour leur 2e portée tandis que les tailles de portées à
la naissance étaient enregistrées pour 4 portées
successives. Les héritabilités du taux d'ovulation (TO),
du nombre d'embryons implantés (EI), de la taille de portée
à la naissance (TP) et du taux de survie prénatal (TP/TO)
étaient respectivement de 0,27 , 0,18 , 0,06 et 0,07. La réponse
à la sélection était de 1,5 ovocytes mais la réponse
corrélée pour la taille de portée était faible
(0,4 lapereau). Ces résultats s'expliquent partiellement par une
réponse corrélée défavorable du taux de survie
prénatal (-0,04). En conclusion il est préférable
de sélectionner directement sur la taille de portée à
la naissance pour améliorer la prolificité.
Moce et al. (Espagne) n'ont pas montré de différences entre
les lignées divergentes sélectionnées sur la capacité
utérine, pour le niveau d'utéroglobuline (une protéine
sécrétée pendant la gestation) et le niveau de progestérone.
Mantovani et al. (Italie) ont montré qu'une évaluation génétique
multi caractères ne présente pas plus d'intérêt
qu'une évaluation génétique mono caractère
pour estimer les valeurs génétiques du nombre de nés
totaux, du nombre de nés vivants et de la mortalité naissance
sevrage. Les corrélations de rang des valeurs génétiques
obtenues avec les 2 méthodes sont en effet très fortes.
Les auteurs notent qu'un BLUP modèle animal classique n'est pas
approprié pour analyser la mortalité naissance sevrage en
raison de la non normalité de la distribution du caractère.
Nagy et al. (Hongrie)
ont démontré la stabilité dans le temps des paramètres
génétiques et des valeurs génétiques calculées
pour le GMQ dans la lignée Pannon sur la période 1999-2007.
Vostry et al. (Hongrie) ont montré l'influence du poids au sevrage
sur le poids à 84 jours pour des lapins de chair HYPLUS issus de
PS59 et de PS119 : les croissances et les indices de consommation ont
été comparés pour 2 groupes constitués en
fonction de leur poids au sevrage (inférieur ou supérieur
à 1300 g). Sur toute la période d'engraissement la différence
de poids entre les 2 groupes reste significative : il n'y a pas de croissance
compensatrice.
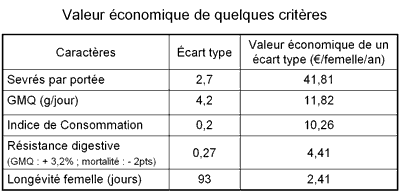
| De façon générale il existe peu de communications relatives aux objectifs de sélection. Le choix des critères de sélection et leurs pondérations relatives sont pourtant déterminants pour définir l'orientation génétique des populations de lapins. Eady et al. (Australie) ont présenté un outil de calcul utilisant des feuilles de calcul du logiciel EXCEL® pour estimer les valeurs économiques relatives des différents critères de sélection. Dans une approche strictement économique ces valeurs peuvent être utilisées pour pondérer les critères d'un objectif de sélection qui combine plusieurs caractères. La méthode repose sur le calcul de la marge brute par femelle et par an à partir de l'ensemble des caractères (taille de portée, mortalité, indice de consommation, poids de vente etc …) et des paramètres économiques (prix de l'aliment, prix de vente, coûts vétérinaires etc ..) qui la composent. La valeur économique relative d'un caractère est l'augmentation relative de la marge brute par femelle et par an résultant de l'augmentation d'une unité de variation (écart-type) de ce caractère. Les caractères de taille de portée au sevrage (45,82 €) et de GMQ (11,82 €) restent les caractères les plus importants d'un point de vue économique. |
 |
Les travaux utilisant les outils moléculaires progressent mais ils sont encore loin de donner lieu à des applications finalisées. 10 communications de génétique moléculaire ont été présentées dans cette session.
Après avoir
mis en évidence l'effet d'un gène majeur agissant sur la
capacité utérine et le nombre d'embryons implantés
dans les lignées sélectionnées de façon divergente
sur la capacité utérine, l'équipe de l'UPV de Valence
a étudié l'influence de deux gènes candidats susceptibles
d'agir sur ces caractères : Le gène codant pour les mélanoprotéinases
matricielles et le gène codant pour l'oviductine. Les mélanoprotéinases
sont des enzymes qui dégradent la matrice extra cellulaire des
tissus conjonctifs et qui semblent jouer un rôle important dans
le développement embryonnaire. Il n'a pas été mis
en évidence d'effet du marqueur moléculaire situé
dans la région du promoteur du gène étudié
sur le taux d'ovulation, le taux de fertilisation à 48 et 72 heures
ou la survie embryonnaire. En revanche le stade de développement
embryonnaire à 72 heures était plus élevé
pour un des génotypes (plus fréquent dans la lignée
à forte capacité utérine). L'oviductine est une protéine
sécrétée par l'oviducte. Elle intervient sur la capacitation
du spermatozoïde. Un effet du génotype de 2 marqueurs situés
dans la région du promoteur du gène de l'oviductine a été
mis en évidence pour la survie embryonnaire mais l'allèle
favorable était plus fréquent dans la lignée à
faible capacité utérine.
Utilisant les mêmes lignées expérimentales, Llobat
et al. (Espagne) ont également étudié l'expression
du gène Oct4 en quantifiant par PCR le niveau de transcription
du gène pour différents génotypes. L'expression du
gène Oct4 au cours du développement embryonnaire a été
mise en évidence chez d'autres mammifères. Il interviendrait
notamment sur la différentiation cellulaire de l'embryon. Le niveau
d'expression du gène n'est pas lié au taux d'ovulation mais
il existe une relation entre le niveau d'expression du gène et
le stade de développement embryonnaire.
En résumé, bien qu'ils apportent des connaissances fondamentales intéressantes, aucun de ces résultats n'est applicable en sélection.
Les communications de Khalil (Arabie Saoudite) et de Ren Keliang (Chine) présentent peu d'intérêt car les méthodologies appliquées sont inadéquates et les résultats présentés sont très discutables.
Le sujet a un intérêt limité pour l'amélioration génétique du lapin de chair mais Fontanesi et al. (Italie) ont le mérite d'avoir identifié des mutations causales (ce qui reste rare dans le domaine de la génétique !) pour 2 gènes intervenant sur la coloration de la fourrure du lapin : les gènes MC1R et ASIP exon 2 codant tous les 2 pour le phénotype Agouti.
Fontanesi et al. ont également étudié 2 gènes candidats susceptibles d'agir sur les caractères de production : le gène codant pour l'hormone de croissance GH et le gène codant pour la myostatine. Aucun polymorphisme n'a été découvert pour le gène codant pour la GH parmi 8 races de lapin. La fréquence d'un allèle du gène de la myostatine est plus élevée chez le lapin nain que chez les races géantes mais il n'a pas été réalisé d'étude de liaison précise entre ce gène et les caractères de croissance.
Deux méthodes de sexage moléculaire ont été présentées (Fontanesi et al. ; Parkanyi et al. (Slovaquie) : les 2 méthodes permettent d'identifier le chromosome Y par PCR.
Le congrès
mondial de cuniculture est traditionnellement une occasion privilégiée
pour décrire des races, des génotypes ou des populations
locales, en particulier pour les pays du Sud. Ces résultats, précieux
pour les filières locales concernées, apportent un intérêt
limité pour les éleveurs français. Ils ne seront
donc que brièvement évoqués.
Les populations locales du Bénin, de la Chine (Rex chinois et race
Saibei) sont respectivement décrites par Akpo et al., Chen Baojiang
et al. et Wu Zhan-fu.
Des expériences de croisement entre populations locales et la lignée
V espagnole ont été réalisées en Egypte (Abou_Khadiga
et al., Iraqui et al.) et en Arabie Saoudite (Al-Saef et al.). En Algérie
une lignée synthétique a été créée
par croisement entre la lignée INRA 2666 (elle-même résultant
du croisement entre la lignée V espagnole et la lignée INRA
2066) et une population locale Algérienne. La création et
le projet de diffusion de cette lignée sont discutés par
Gacem et al. Ces travaux ont pour but de combiner les qualités
d'adaptation des populations locales (résistance à la chaleur,
robustesse) et les qualités de productivité (prolificité
et croissance) des lignées européennes.
Les performances de races classiquement utilisées en Europe (Néo
Zélandais, Californien, Géant Blanc du Bouscat) élevées
dans les conditions locales en pures (Hongrie) ou en croisement (Québec)
sont décrites respectivement par Ouyed et al. et Nofal et al.