![]()
5 février 2009 - Journée d'étude ASFC « Vérone
- Ombres & Lumières »
Travaux
de la session "Comportement et Bien-Être", du
9ème Congrès Mondial de Cuniculture
par
Gérard COUREAUD * et Anouk DRONNEAU **
* Centre Européen des Sciences du Goût, UMR 5170 CNRS/UB/INRA
- 21000 Dijon
** Coopérative des Producteurs de Lapins du Bocage, 85700 Réaumur

A. Dronneau lors de la présentation
La session de Bien-être
et Comportement, présidée par Mme Gabriela Gonzalez-Mariscal
(Mexique), a permis la présentation de 26 communications (dont
1 présentation invitée et 25 communications courtes), soit
une augmentation de plus de 50% par rapport au précédent
Congrès mondial (12 présentations dont 1 conférence
invitée - rappelons que cet avant dernier Congrès avait
eu lieu hors Europe).
Sur les 26 communications, 11 avaient une orientation plus singulièrement
comportementale, les 15 autres plus bien-être.
La provenance des communications était la suivante:
Europe: 23 - Afrique: 1 (Nigeria) - Amérique du Nord: 1 (USA) -
Amérique du Sud: 1 (Brésil)
En Europe, la sous-répartition était la suivante:
Italie: 11 - Hongrie: 5 - Espagne: 2 - Allemagne: 2 - France: 1 - Pays-Bas:
1 - Pologne: 1
Rapport de synthèse invité

G. Coureaud au cours de la présentation du rapport de synthèse
pendant le Congrès à Vérone en juin 2008
La conférence invitée de Coureaud et al. (pages 317-338 des Proceedings du Congrès) traite du développement des capacités sensorielles, cognitives et comportementales du lapereau, entre la naissance et le sevrage. Elle illustre ce développement au travers de travaux de la littérature et au travers de ceux menés par les auteurs au cours des 10 dernières années. Elle intègre notamment les études menées ou supervisées par les éthologistes dijonnais du CESG sur l'importance des odeurs dans la relation mère-jeunes, donc des informations olfactives impliquées dans l'interaction vitale du jeune avec l'élément crucial de l'environnement social et alimentaire que représente la mère. Une description des relations mère-lapereaux à l'état semi-naturel est également faite. Elle est intéressante dans le sens où un des principes du bien-être pour le " Farm Animal Welfare Council " est la possibilité pour l'animal d'exprimer le comportement normal de l'espèce. On peut ainsi noter que dans les conditions domestiques, la femelle a accès à un nid isolé, peut gratter la litière et y rajouter du poil, ce qui ressemble au comportement naturel d'aménagement d'un terrier. La femelle domestique ne peut par contre pas refermer son nid à la fin de l'allaitement. Les lapereaux ont donc l'occasion de solliciter la femelle en dehors des heures de tétées, à partir de 13 jours d'âge environ, c'est-à-dire lorsqu'ils parviennent à sortir de la boîte à nid. C'est pourquoi des systèmes de logement avec plateforme ont été imaginés pour permettre à la lapine de s'isoler de sa portée. Mais, dans ces systèmes, des données (non publiées pour l'instant) montrent que ces tentatives deviennent malgré tout fréquentes dans les 2 heures (3ème semaine) à 30 minutes (5ème semaine) avant le moment prévu de l'allaitement ; la stratégie de l'installation de plateformes paraît donc discutable car les interactions des jeunes avec la mère semblent plus nombreuses, les lapereaux étant stimulés par le retour de la lapine en partie basse de la cage (Mirabito et al., 2004).
Au vu des observations faites par Coureaud et al. vis-à-vis des communications olfactives entre les mères et les jeunes, deux questions pourraient notamment se poser. Peut-on optimiser cette relation, et par là améliorer la qualité de l'allaitement ? Saisi-t-on toujours l'impact, si il y en a un (ce qui semble le cas ; cf. Coureaud et al., 2002), d'un changement d'aliment entre la période de gestation et celle de l'allaitement par rapport à une éventuelle difficulté que pourrait rencontrer le lapereau dans sa reconnaissance de la mère ? Les travaux menés à Dijon sur cette thématique vont se poursuivre au cours des années à venir, afin notamment de préciser si les odeurs biologiquement émises par la mère jouent un rôle direct sur la survie, la croissance et le bien-être des animaux en période pré- et péri-sevrage.
Résumés/Commentaires des communications libres.
Notons qu'en dehors de la conférence invitée, la grande majorité des travaux à angle d'approche majoritairement comportementale ont porté sur les animaux en engraissement. Seule les études de Zucca et al. et de Gerencser et al. concernent directement la période de pré-sevrage, la première plus côté comportement du lapereau, la seconde côté comportement de la femelle allaitante.

Lapins Néo-Zélandais Blanc et Hollandais (Dutch belted)
Le travail de Zucca et al. (Italie) s'est penché sur l'effet potentiel d'une manipulation précoce de l'animal par l'Homme (" handling ") sur la diminution de la réponse de peur envers un nouvel environnement et envers l'Homme (ce travail s'inscrit dans la continuité de travaux de la littérature ; ex. Wyly et al., 1975). Il suggère qu'une exposition minime à un contact humain (main descendue dans le nid au-dessus de la portée) en phase de pré-tétée chez le nouveau-né réduit le temps d'émergence et d'immobilité dans des épreuves comportementales d'évaluation du stress. Cela mériterait sans doute réplication et étude sur un effectif plus large pour consolider cet effet potentiellement très intéressant. Rappelons qu'en pratique, les nids sont visités par l'éleveur au minimum 2 à 3 fois sur la première semaine après mise-bas, mais après la tétée. Il serait intéressant de savoir si ces manipulations post-tétée ont le même impact que celles décrites dans l'étude. Notons que peut-être rapproché, dans une certaine mesure, de cette communication, celle de Pritt et al. sur la socialisation du lapin Dutch belted (DB). Ce lapin beaucoup utilisé aux USA présente une taille plus petite (d'environ 2/3) que le Néo-Zélandais (NZ) et pourrait de fait être plus facile de manipulation en tant qu'animal de laboratoire. Le problème est qu'il est spontanément plus agressif et moins adaptable à un changement d'environnement que le NZ. Les auteurs ont donc initié un programme de socialisation visant à faire s'habituer une colonie de DB à des manipulations humaines répétées en périodes de pré- et post-sevrage, à des enrichissements de l'environnement, et autres procédures (non précisées) classiquement utilisées chez le NZ. Le programme, préliminaire, annonce un succès relatif du fait d'un plus faible nombre de remarques pointant un comportement difficile des animaux, de la part des utilisateurs de lapins issus de ce programme. Une description de ce programme de socialisation, avec une hiérarchisation des résultats, aurait été appréciée.
L'étude de Gerencsér et al. (Hongrie) a porté quant à elle sur l'impact du régime lumineux sur la fréquence et la durée d'allaitement (lapin Pannon White). L'idée était de maintenir les femelles d'un groupe sous un régime lumière/obscurité constant de 16h/8h sur la période pré- puis post-parturition, alors que dans l'autre groupe les femelles basculaient en régime 8h/16h dans les jours qui suivaient la mise-bas (sauf pour quelques jours autour de l'insémination). Le fait de modifier le régime lumineux a pour effet d'augmenter la fréquence apparente des allaitements multiples (il n'est pas vérifié que les visites supplémentaires au nid se traduisent par de réels allaitements) et de réduire la durée moyenne de chaque allaitement. Soulignons que la recommandation au niveau des élevages en terme d'intensité lumineuse est de 100 lux., contre 40 à 70 dans cette étude. Aucun critère technique n'a été exploré, ni les conséquences de ce genre de programme après répétitions. Aucune visée appliquée de ce type de modification n'est suggérée en termes de pratique d'élevage.
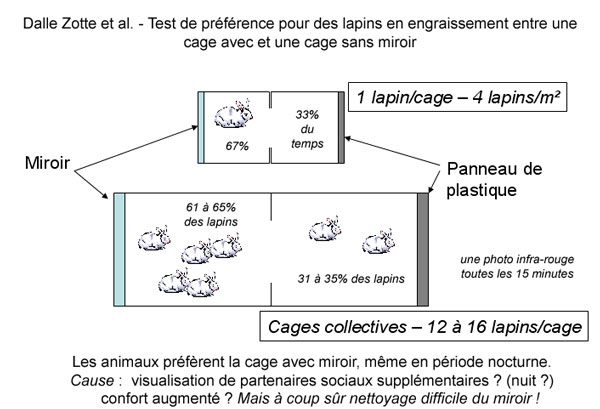
Dans un esprit différent, plusieurs études se sont intéressées à des aspects d'enrichissement de l'environnement. Ainsi, Dalle Zotte et al. (Italie) ont évalué la préférence de lapins en engraissement pour des cages individuelles ou collectives disposant ou non de miroir. L'hypothèse semblait être que l'ajout de miroirs dans les cages constituerait un facteur d'enrichissement bénéfique au bien-être de l'animal, bien qu'il ne soit pas dit pour quelles raisons (visualisation de partenaires sociaux supplémentaires ? Effet de groupe " sécurisant " ?). Les animaux semblent préférer (en situation de choix entre cages avec et sans miroirs) celles possédant un miroir. Assez curieusement, cette préférence s'exprime même au cours de la période nocturne, donc à un moment où l'animal ne peut voir de reflet dans le miroir. Cela peut laisser penser que l'avantage conféré par le miroir n'est pas directement (ou uniquement) lié à un enrichissement visuel, mais peut-être à un isolement social et thermique augmentant le confort. Ce genre d'étude a un intérêt scientifique pur, mais parait très difficilement applicable en élevage du fait de la sécurité de l'éleveur et de l'utilisation de nettoyeurs Haute Pression.
Dans un esprit proche, Negretti et al. (Italie) se sont interrogés sur la recherche d'informations visuelles homospécifiques (donc d'interactions sociales) par l'animal en cage, en quantifiant l'orientation préférentielle du regard d'individus introduits dans un système à 3 cages adjacentes (une centrale, deux latérales). Les auteurs concluent que les animaux tendent à se regarder les uns les autres et donc à établir une interaction sociale par le biais de cette modalité sensorielle, dès lors qu'au moins un autre individu est présent dans une cage adjacente, ce qui suggère in fine que la disposition classique des animaux dans les batteries d'élevage est judicieuse car conservatrice de ce besoin de l'animal. Plusieurs éléments méthodologiques manquent néanmoins à ce stade pour aboutir à une conclusion sereine (mesure discutable de l'orientation réelle du regard, absences de groupes contrôles).
D'autres observations ont été faites sur l'enrichissement de l'environnement chez les lapins en engraissement :
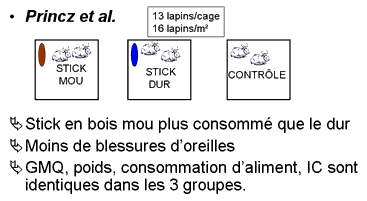
Princz et al. (Hongrie) ont comparé 3 groupes de lapins : mise à disposition de stick durs ou mous et un 3ème groupe de contrôle. Les critères zootechniques (GMQ, poids, consommation d'aliment, IC) sont comparables dans les 3 groupes. Le stick est plus consommé quand le matériau est mou. La fréquence d'observation de blessures d'oreilles est moindres quand des " jouets " sont disponibles (16 lapins/m² en parc de 13 lapins abattus à 11 semaines d'âge).
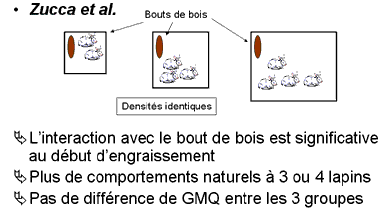
Zucca et al (Italie) ont installé des groupes de 2, 3 ou 4 lapins/cage (à densité identique). Certaines cages étaient enrichies avec un bout de bois. L'interaction avec le bout de bois est significative au début de la période d'engraissement et liée avec une plus grande fréquence de toilettage. Les cages de 3 ou 4 lapins exprimaient plus de comportements naturels de déplacement et de posture que par 2. Il n'y avait aucune différence de GMQ sur les différents groupes.
 |
 |
Rizzy et Chiericato (Italie) ont étudié des lapins mâles de la souche Hycole logés en cages individuelles de 40 à 80 jours. Deux photopériodes ont été étudiées (9 et 5 heures de lumière) et certaines cages étaient enrichies avec un bout de bois. Le temps d'éclairement n'a pas influencé les performances de production, mais les lapins du groupe " bout de bois + nourriture " présentaient une ingestion et un GMQ supérieur. La photopériode plus courte diminuait par contre le taux de calcium dans les os (impact négatif).
Au final, les conclusions concernant les performances zootechniques vis-à-vis d'un enrichissement des cages par un objet (par exemple bout de bois) sont parfois contradictoires, mais les différences avec les groupes témoins sont rarement observées. D'après d'autres sources (Mirabito et al. 2000), de la mortalité est néanmoins observée dans certaines cages disposant de bouts de bois ajoutés, suggérant une contamination par voie orale des animaux. Par contre, l'intérêt de ces enrichissements sur le bien-être des animaux se partage de manière plus unanime.
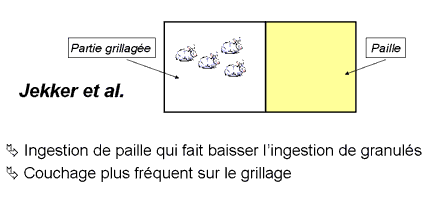
Jekkel et al. (Hongrie) ont conduit un essai visant à déterminer le comportement de lapins Pannon (prise alimentaire, locomotion, repos, comportements autodirigés et agonistiques) dans des cages collectives d'engraissement d'une hauteur de 80 cm dont le fond était recouvert, ou pas, d'une épaisse couche de litière de façon plus ou moins prolongée entre 5 et 11 semaines. L'analyse des comportements souligne que le principal effet engendré par l'ajout de litière est un effet alimentaire: les animaux consomment cette dernière de façon importante, ce qui se traduit par une réduction de l'ingestion de granulés. L'intérêt d'un tel ajout pour le bien-être ne paraît donc pas évident (même si l'analyse comportementale conduite ici demeure relativement succincte), d'autant que le couchage est plus fréquent sur le grillage que sur la paille qui se salit au long de la période d'engraissement.
 |
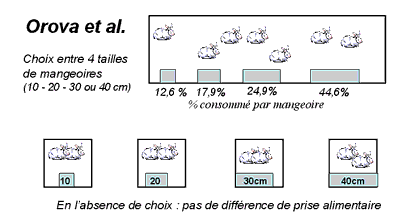 |
D'approche également comportementale, il faut noter le travail intéressant de Kowalska et al. (Pologne), portant sur une tentative de sélection de lapins Néo-Zélandais sur la base d'un trait comportemental lié à la réaction au stress. L'état émotionnel de mâles et de femelles nullipares a été quantifié par des mesures comportementales (réaction à l'introduction dans un open-field, mesure de l'immobilité tonique) ou physiologique (hyperthermie), et ces données ont été analysées en lien avec le succès reproducteur lors de la 1ère saillie et mise-bas. Les tests comportementaux ont permis d'isoler des individus dits " actifs " (A; explorateurs, à priori peu stressés) et des individus " passifs " (P; à priori plus stressés). Les mâles A sont plus nombreux (80%) à copuler que les P (20%). A l'inverse, 70% des femelles P sont saillies contre 45% des A. Chez les femelles saillies, la mise-bas a essentiellement lieu dans la noite à nid chez les P, alors que les A tendent à mettre bas plus fréquemment dans la cage. Le taux de mortinatalité est plus élevée chez les A (22,2%), dont les portées sont par ailleurs plus petites (5,2 contre 6,8 pour les P). Les auteurs précisent que le comportement et les soins maternels sont plus prononcés chez les P, mais il n'est pas précisé comment cela est quantifié. Le profil " passif " semble donc plus adapté à la vie en confinement dans les cages de dimensions standards.
Enfin, de façon
intéressante, 3 études se sont penchées sur le bien-être
des animaux au cours de la période de transport jusqu'à
l'abattoir, ou lors de l'abattage lui-même.
Vignola et al. (Italie) ont tenté de déterminer si le mode
de transfert de la salle d'élevage jusque dans le camion de transport
vers l'abattoir (mode " doux " ou " agité ",
donc à priori respectivement peu ou très stressant), et
si la position de l'animal dans le camion (caisses du haut versus du bas,
à l'arrière ou à l'avant, donc offrant des conditions
de température et d'humidité contrastées) influençait
le niveau de stress et l'état corporel de l'animal. Si les lapins
transportés dans les cages hautes et situées à l'arrière
semblent présenter des signes de déshydratation (température
plus élevée), les effets stressants occasionnés par
le transport, semblent plus liés au transport lui-même et
à la manipulation des animaux, qu'au mode de transfert de l'élevage
au camion et à la position dans ce dernier (pas de différence
entre les groupes).
Nodari et al. (Italie) ont suivi 1020 animaux au moment de leur abattage
en Italie (abattage classique par électronarcose suivi d'une saignée).
Il est apparu que dans l'abattoir où a eu lieu l'étude,
dans 10,8% des cas le système d'électronarcose n'était
pas correctement appliqué, et que 3 lapins (0,3%) étaient
encore conscients au terme de cette dernière. Plus généralement,
18 animaux (1,8%) se sont " réveillés " partiellement,
l'indicateur le plus fiable de ce réveil semblant être le
réflexe cornéen. Au vu de la cadence d'abattage, cette méthode
ne peut pas servir comme moyen de contrôle pour les techniciens
sur chaîne. Quels sont les seuils d'échec acceptables vis-à-vis
d'une méthode ?
Lopez et al. (Espagne) ont abordé la question du bien-être
de l'animal au cours de l'abattage, et de la qualité musculaire
de la viande à son issue, au cours d'abattages conduits selon la
tradition Halal, en comparant les conséquences de ce type d'abattage
avec celles de l'abattage par électronarcose préalable.
Le suivi d'un petit nombre d'animaux (comparativement à l'étude
de Nodari et al. notament) - 30 dans le groupe standard, 20 dans le groupe
Halal - souligne une absence de vocalisations, spasmes et autres mouvements
marqués au cours de l'exsanguination selon la méthode Halal,
et un relâchement habituel du corps lors de la suspension sur la
chaîne. Ni hémorragie ni endommagement osseux ne sont observés.
Si certaines variations existent en terme de couleur de la carcasse et
des muscles, au moins ponctuellement, elles ne semblent pas jouer sur
la qualité de la viande en tant que telle. Les deux méthodes
aboutissent donc à des observations dans les normes des procédures
d'abattage de l'espèce.