| |
Introduction
: les os principaux Les
os du squelette | |
Pour
une description complète du squelette et de la musculature, il est conseillé
de consulter les ouvrages spécialisés comme "l'Atlas d'Anatomie
du Lapin" publié par Barone et al. (1973).
Par ailleurs, un article
comme celui de Cantier et al. (1974) donne une correspondance entre les dénominations
des muscles squelettiques adoptées par différents auteurs.
Les principaux
os du squelette sont illustrés sur la figure 5 ci-contre (cliquer
sur l'image pour l'agrandir) |
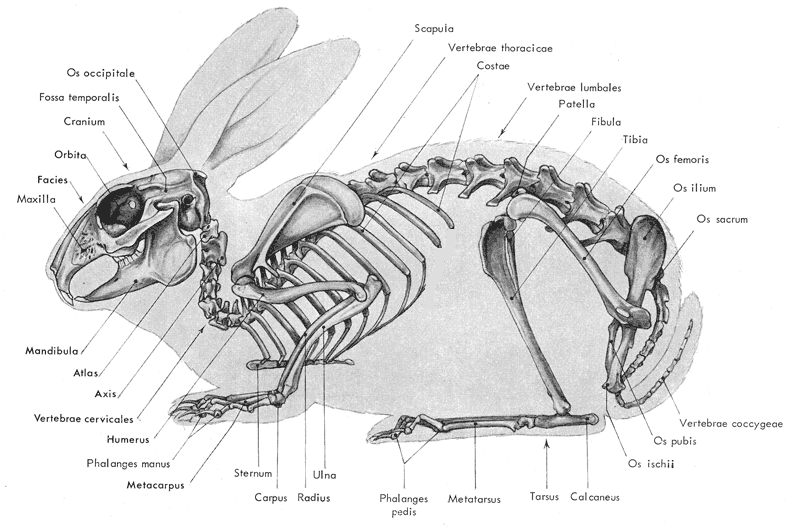
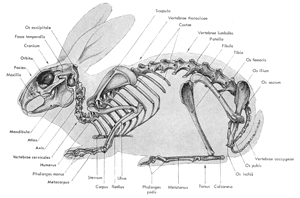
Figure
5: Schéma du squelette d'un lapin
(d'après
Barone et al., 1973)
| |
| |
Le crâne et la
dentition Retour
haut de page  | |
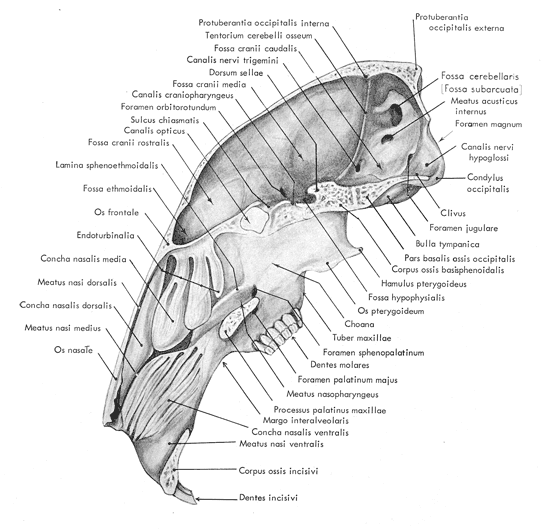
| La
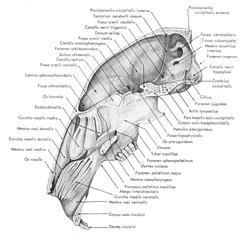
figure 6 correspond à une section médiane du crâne
dans le sens longitudinal. Il convient d'y remarquer l'ampleur des sinus nasaux
qui occupent près du tiers du volume intérieur du crâne. Une
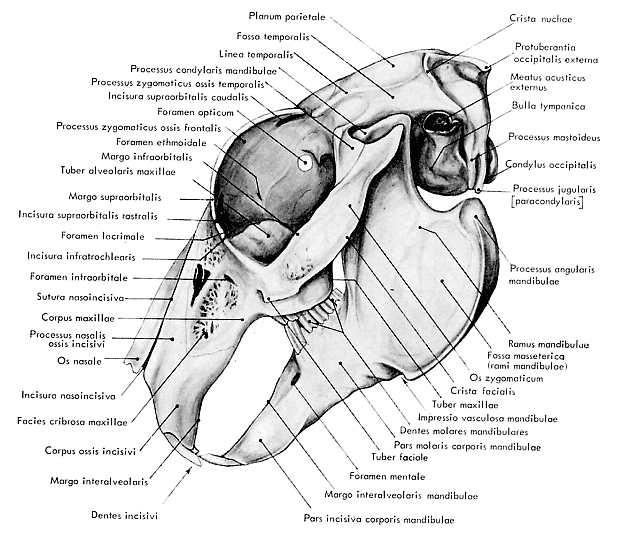
vue latérale du crâne est illustrée par la figure 7
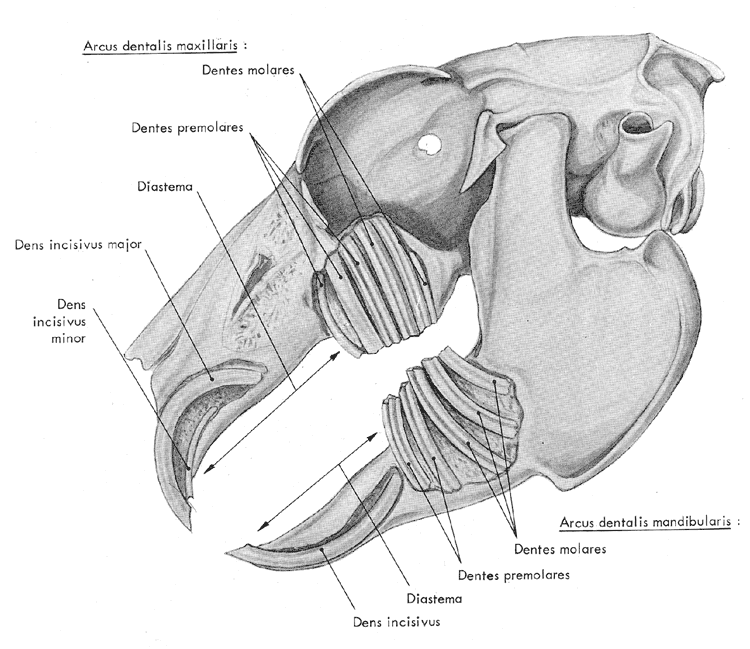
L'implantation
des dents est illustrée sur la figure 8. Le lapin
possède 2 paires d'incisives à la mâchoire
supérieure et une seule à la mâchoire inférieure.
Ceci a conduit très tôt les zoologistes à
distinguer d'une part les lapins (et les lagomorphes en général)
et d'autre part les rongeurs qui n'ont qu'une seule paire d'incisives
à chaque mâchoire. Chez le lapin, la deuxième
paire se place derrière la première qui la cache
totalement. Les incisives sont entièrement revêtues
d'une couche d'émail qui est plus mince en arrière
qu'en avant; ceci permet au lapin d'affûter ses dents en
biseaux en usant celles du haut contre celles du bas et réciproquement.
Leur face antérieure porte un sillon longitudinal. Il n'y
a pas de canines et un fort diastème sépare les
incisives des prémolaires ( 3 + 2
paires) et des molaires ( 3 + 3 paires)
|
Figure 6: Coupe médiane
du crâne d'un lapin
(d'après Barone et al., 1973)
cliquer
sur l'image pour l'obtenir plein écran
(ou ici  pour une image encore 2 fois plus grande et donc plus lisible, mais dépassant
un simple écran)
pour une image encore 2 fois plus grande et donc plus lisible, mais dépassant
un simple écran) | |
| | | |
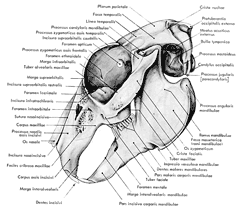
Figure
7 : Vue latérale externe du crâne du lapin
(d'après Barone et al., 1973)
| Figure
8 : Vue de l'implantation des dents chez le lapin. L'os a été
dégagé pour laisser apparaître la base des dents (d'après
Barone et al., 1973)
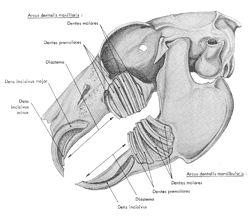
| |
| | | |
Pour l'ensemble
de la mâchoire, à la partie supérieure il
y a donc deux paires d'incisives, 3 paires de prémolaires et
3 paires molaires. A la partie inférieure il y a 1 paire d'incisives,
2 paires de prémolaires et 3 paires de molaires, soit 28 dents
en tout dont 26 seulement ont un rôle fonctionnel. Comme les dents
de tous les lagomorphes, celles du lapin sont profondément insérées
dans les mâchoires mais sans racine (figure 8). En effet, la
croissance de toutes les dents est continue, durant toute la vie
de l'animal. Une mesure de la vitesse de croissance des incisives a
donné une valeur de 2 mm par semaine pour la mâchoire supérieure
et de 2,4 mm pour la mâchoire inférieure. Celle des molaires
et prémolaires (dites aussi dents "jugales") est beaucoup
moins rapide et a été estimée à 2 mm par
mois seulement (~ 4 fois moins rapide)
Enfin, signalons que la dentition de lait présente à
la naissance des lapereaux (incisives et prémolaires) tombe aux
environs du 18e jour après la naissance et est presque immédiatement
remplacée par la dentition définitive.
|
| | Les
vertèbres et les os des membres
Retour
haut de page
| |
Au plan squelettique,
l'ossature du lapin comprend 7 vertèbres cervicales, dont
les deux premières sont l'atlas (portant la tête) et l'axis
qui joue un rôle primordial dans la rotation de la tête. Les
12 vertèbres thoraciques portent les 12 côtes.
Seules les 10 premières côtes sont reliées ventralement
au sternum pour former la cage thoracique, les 2 dernières sont
dites "flottantes". Les 7 vertèbres lombaires
sont suivies par le sacrum. Ce dernier correspond à la fusion
de 4 vertèbres sacrées. Il porte les os du bassin. Il est
suivi par 15 vertèbres coccygiennes dont la dernière
dizaine correspond à la queue du lapin.
Dans le membre antérieur,
le radius et le cubitus (appelé aussi ulna, le plus long des 2 os de l'avant-bras)
sont au contact l'un de l'autre mais sans fusion. Dans le membre postérieur,
tibia et péroné ont par contre presque totalement fusionné
dans leur partie distale. La partie libre de ce qui "reste" du péroné,
est appelé fibule.
|
| | Structure
générale des os et croissance osseuse | | Les
os présentent deux aspects structuraux bien distincts. A la périphérie,
une "couche" dure et dense, c'est l'os compact. Au centre, creusé
de petites cavités, c'est l'os spongieux. Compact ou spongieux, le tissu
osseux adulte est de type lamellaire. L'architecture de l'os compact s'appuie
sur une belle ordonnance de lamelles concentriques. Dans l'os spongieux, les lamelles
osseuses forment un treillis de travées minces, irrégulière.
A l'intérieur des os longs se trouve la moelle osseuse responsable de la
fabrication des éléments figurés du sang et du système
immunitaire : globules rouges, globules blancs et plaquettes.
|
| |
Les cellules de l'os
Ostéocytes
Ostéoblastes
Ostéoclastes
|
| Trois
types de cellules participent à la vie et au métabolisme du
tissu osseux : les ostéocytes, les ostéoblastes, les ostéoclastes.
Un réseau de vaisseaux sanguins assure leur alimentation.
Les ostéocytes sont des cellules emprisonnées dans la matrice
osseuse mais qui communiquent entre elles par de fins prolongements connectés
ensemble. Ces cellules sont des " mécano-récepteurs "
sensibles aux variations de pression qui s'exercent sur la matrice osseuse.
Les ostéoblastes sont des cellules situées à
la surface des travées osseuses. Leur rôle est de synthétiser
les éléments de la matrice osseuse et de permettre sa calcification.
Ces cellules communiquent entre elles et avec les ostéocytes de la matrice
grâce à des systèmes de jonctions cellulaires spécialisées.
Les ostéoclastes sont des cellules possédant plusieurs
noyaux. Elles résorbent localement la surface des travées osseuses
sous forme de petites érosions à l'origine d'une zone ou encoche
de résorption. Ces cellules ne sont pas reliées aux autres cellules
osseuses par des jonctions mais sont couplées par des médiateurs
solubles qui circulent d'une cellule à l'autre.
|
| | La
croissance du squelette | | Le
développement du squelette est initialement assuré par les points
d'ossification (à partir de membranes fibreuses de cartilage). Ensuite,
la croissance des os en diamètre et en épaisseur est assurée
par le travail combiné des ostéoblastes et des ostéoclastes
qui renouvellent continuellement la masse osseuse, avec une dominance de l'activité
des ostéoblastes pendant la période de croissance. La croissance
en longueur est assurée à partir du cartilage de conjugaison situé
à la base de l'épiphyse de chaque os (ou plaque épiphysaire).
La croissance en longueur est terminée, donc la taille du lapin est fixée,
vers 140 à 150 jours lorsque la plaque épiphysaire est "fermée",
c'est à dire qu'il y a disparition du cartilage de conjugaison, en grande
partie sous l'action des hormones stéroïdes.
|
| | Muscles
et croissance musculaire
Retour
haut de page
| |
Les masses musculaires quantitativement
les plus importantes sont celles de la partie arrière du corps du lapin
: râble et cuisses. Les muscles squelettiques ont pour fonction d'assurer
les mouvements de l'animal. Ils ont donc 2 ou 3 points de fixation au squelette
pour que leur contraction puisse engendrer un mouvement.
Du point de vue "boucher",
le muscle ayant la masse la plus importante est le muscle longissimus dorsi.
Au plan anatomique cette masse musculaire correspond à une succession de
muscles différents, le long dorsal, le long épineux, et le muscle
multifide dorsolombaire (longissimus lumborum). Très
divers quant à leur forme, leur taille et leur fonction, les muscles du
squelette se caractérisent aussi par une forte hétérogénéité
tissulaire. Le tissu nerveux (fibres motrices afférentes et fibres sensitives
efférentes) assure le contrôle de la contraction et de la relaxation
musculaire. Les vaisseaux sanguins assurent la fourniture des nutriments et de
l'oxygène, et évacuent les produits du catabolisme musculaire. Les
fibres musculaires constituent 75 à 90% du volume musculaire et représentent
l'élément de base du muscle strié squelettique. Les fibres
musculaires sont entourées d'une mince gaine de tissu conjonctif riche
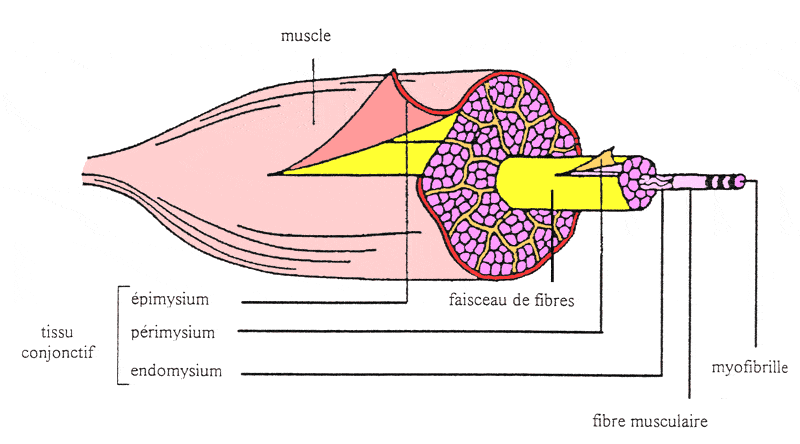
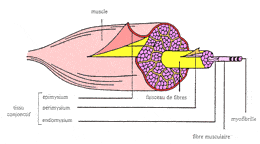
en collagène, l'endomysium (figure 9). |
| | | | Figure
9 : Structure schématique d'un muscle (d'après
Gondret, 1997)
 |
| | Les
fibres musculaires | | Les
fibres musculaires sont réunies en faisceaux primaires enveloppés
d'une gaine conjonctive fine, le périmysium. Entre ces faisceaux
primaires peuvent se trouver des cellules adipeuses appelées adipocytes
intracellulaires. Les faisceaux primaires sont à leur tour tous regroupés
en faisceaux secondaires enveloppés dans une gaine de périmysium
plus épaisse. Enfin, l'épimysium regroupe tous les gros faisceaux
secondaires pour former l'enveloppe du muscle lui-même.
Aux
extrémités du muscle, cette trame conjonctive forme les tendons
et les aponévroses assurant ainsi la liaison du muscle avec le squelette.
Enfin, le tissu aréolaire situé dans les espaces entre les
fibres musculaires, forme un réseau intriqué prenant l'aspect d'un
grillage élastique plus ou moins développé. Son rôle
est de munir le muscle d'une charpente organisant les fibres musculaires, d'empaqueter
les éléments nerveux et vasculaires et de présenter des surfaces
lubrifiées nécessaires aux mouvements des fibres musculaires entres
elles et des faisceaux entre eux. |
| |
Les fibres musculaires
et myofibrilles | | La
fibre musculaire est une cellule allongée, tubulaire et plurinuclée.
Elle constitue l'unité de base de l'activité motrice. La plus grande
part du sarcoplasme de la cellule musculaire est occupée par des éléments
contractiles : les myofibrilles.
Il existe 3 types des filaments distincts dans les myofibrilles. Les filaments
épais (150 Å de diamètre) sont essentiellement composés
de myosine. Les filaments fins (70 Å de diamètre) sont composés
essentiellement d'actine. Ils contiennent également de la tropomyosine
et le complexe troponine. Les filaments intermédiaires constituent un réseau
de filaments longitudinaux et transversaux associés au sarcomère.
Lorsque les faisceaux de fibres vont d'un bout à l'autre du muscle avec
une direction parallèle à l'axe principal, le muscle est dit "fusiforme"
(par exemple biceps femoris). Par contre lorsque les faisceaux forment
un angle plus ou moins marqué par rapport à l'axe principal, le
muscle est dit "penné" (exemple longissimus lumborum). |
| | Retour
haut de page | | La
myosine et l'actine jouent un rôle central architectural et un
rôle enzymatique clé dans la contraction musculaire au cours de laquelle
l'énergie chimique (ATP) est transformée en énergie mécanique.
La contraction musculaire correspond ainsi à un glissement des filaments
fins d'actine entre les filaments épais de myosine. L'énergie nécessaire
à la contraction provient de l'hydrolyse de l'ATP par l'ATPase myofibrillaire.
Cet ATP est ensuite régénéré par un métabolisme
aérobie ou anaérobie. En fonction de la vitesse de contraction des
fibres au sein d'un muscle et du type majoritaire du métabolisme mis en
œuvre pour régénérer l'ATP utilisé lors de la
contraction, on a pu distinguer différents types de fibres. Les
caractéristiques fonctionnelles des différents types de fibres sont
résumées au tableau 3. Compte tenu de la teneur très variable
en myoglobine d'un type de fibre à l'autre et du fait que certains muscles
sont constitués quasi uniquement de fibres d'un type donné, le lapin
possède des muscles de couleur rouge ou complètement blanc.
|
| | Les
principaux types de fibres musculaires et leurs caractéristiques | | Tableau
3 : Caractéristiques des différents types de fibres musculaires
(d'après
Gondret, 1997)
| Caractères |
Types
de Fibres | | I |
IIA |
IIX |
IIB |
| Vitesse
de contraction | lente |
rapide |
rapide |
rapide |
| Résistance
à la fatigue | +++ |
++ |
? |
? |
|
|
| Surface
de la plaque motrice | + |
+++ |
? |
+++ |
| Réticulum
sacoplasmique | +++ |
+++ |
? |
+++ |
| Tubules
transverses | + |
+++ |
? |
+++ |
| Couleur |
rouge |
rouge |
blanc |
blanc |
| Myoglobine |
+++ |
+++ |
++ |
+ |
| Densité
des capillaires | +++ |
++ |
? |
+ |
| Nombre
de mitochondries | +++ |
+++ |
++ |
+ |
| Richesse
en collagène | +++ |
++ |
? |
++ |
| Aire
de section | + |
++ |
+++ |
++++ |
| | |
Utilisation
du glycogène | + |
++ |
++ |
++++ |
| Utilisation
des lipides | +++ |
+++ |
? |
+ |
| |
| ATPase
myofibrillaire | +++ |
++ |
? |
+ |
| Hexokinase |
+++ |
++ |
? |
+ |
| Phosphorylase |
+ |
++ |
? |
++ |
| Enzymes
de la glycolyse anaérobie |
+ |
++ |
++ |
++ |
| Enzymes
oxydatives (aérobies) |
+++ |
++ |
++ |
+ |
|
| | | | |
| | | |
Biologie
du Lapin - Fin du chapitre 3 " Squelette
et croissance musculaire" |
| | | |
|
| | | |
|
| | | | |