| |
|
1- Organisation
de la Sélection
La
sélection de lignées destinées à être
utilisées sans croisement pour la production de lapins de
chair est tout à fait possible, mais présente quelques
inconvénients et risques
|
|
Les principales
caractéristiques des schémas d'amélioration
génétique du lapin ont été présentées
par BASELGA [1], dans son rapport
invité en se référant au croisement 3 voies
pour la production du lapin de chair. Cette organisation de la sélection
est la plus répandue. Elle repose sur l'utilisation de lignées
spécialisées mâles et femelles. GARREAU et
al. [2] dans le second rapport
invité rappellent qu'une méthode alternative moins
coûteuse consiste à utiliser une seule souche sélectionnée
conjointement pour la prolificité et la croissance. Des souches
ont ainsi été sélectionnées avec succès
pour un objectif global en France, en Espagne, au Brésil
et en Australie. Cependant l'utilisation d'une seule souche ne permet
pas de bénéficier de l'effet d'hétérosis
et accroît les conséquences néfastes d'une réduction
de la variabilité génétique dans les populations
sélectionnées.
Les critères
retenus pour la création de nouvelles souches ont été
discutés : Une méthode consiste à appliquer
de très fortes intensités de sélection en choisissant
les reproducteurs de la nouvelle souche au sein d'une population
de base très large, issue, par exemple, de plusieurs élevages
de sélection ou de multiplication. Les lignées "
hyperprolifiques " et " hyperlongèves "
(animaux ayant une très forte prolificité ou une très
grande durée de vie productive) sélectionnéesà
l'Université de Valence (Espagne) ont été créées
en appliquant ce principe. On peut toutefois s'interroger sur la
pertinence des critères de sélection (plus de 20 portées
pour les femelles de la lignée hyperlongève) et sur
la validité de l'estimation de la valeur génétique
de ces animaux élevés dans des milieux hétérogènes.
Les lignées
dites "mâles" sont principalement sélectionnées
sur la vitesse de croissance post sevrage ou sur le poids à
l'âge d'abattage. Les lignées dites "femelles"
sont généralement sélectionnées sur
la taille de portée à la naissance ou au sevrage,
mais l'amélioration de l'aptitude des femelles à élever
leurs lapereaux apparaît aujourd'hui dans certains schémas
par la prise en compte des poids individuel ou des poids de portées
au sevrage.
|
|
La sélection
sur la vitesse de croissance donne rapidement des résultats
: + 1 à +3 % de GMQ, par génération.
Celle sur la
prolificité est aussi efficace mais un peu moins rapide :
+0,6 à +1,4% de lapereaux par mise bas et par génération
|
La méthodologie
d'estimation des valeurs génétiques la plus courante
est celle du BLUP. Les estimations de progrès génétique
rapportées dans la littérature sont comprises entre
18 et 35 grammes/génération pour le poids à
l'abattage et comprises entre 0,45 et 1,23 g/jour/génération
pour la vitesse de croissance. Les réponses corrélées
à cette sélection sont une augmentation du format
adulte, de la masse du tube digestif et des quantités d'aliment
ingérées. La sélection pour la croissance a
peu d'incidence sur l'indice de consommation et le rendement de
carcasse. Les estimations du progrès génétique
en lignée femelle sont comprises entre 0,05 et 0,13 lapereaux
nés vivants ou sevrés par portée née
|
| |
Selon
les lignées, cet accroissement de la prolificité résulte
principalement soit de l'augmentation du taux d'ovulation, soit d'une
amélioration de la survie embryonnaire. La réponse à
la sélection mesurée sur les femelles croisées
est légèrement supérieure à celle mesurée
sur les souches pures. En revanche la réponse à la sélection
mesurée sur les issus du croisement terminal est plus faible,
probablement en raison d'une interaction entre l'aliment distribué
dans les élevages de production (variable d'un
élevage à l'autre) et le niveau génétique
des caractères de croissance.
Enfin deux approches
destinées à améliorer la diffusion du progrès
génétique vers les étages de production ont
été présentées. En France les sélectionneurs
liés à l'INRA par des contrats de partenariat ont
créés un étage supplémentaire, appelé
étage de démultiplication, entre le noyau de sélection
et l'étage de multiplication. Les élevages de "
démultiplication " sélectionnent les femelles
de la même façon que dans le noyau de sélection
et accouplent ces femelles avec des mâles provenant du noyau
de sélection. En Espagne, les élevages de multiplication
(coopératives, sociétés privées de sélection)
sont intégrés à l'opération de sélection.
Chacun de ces multiplicateurs entretient et sélectionne des
noyaux secondaires de souches femelles pures et accouplent ces femelles
avec les mâles de la souche femelle réciproque, issus
du noyau primaire, pour produire les femelles croisées mères
du lapereau de boucherie.
|
|
2 - Variabilité
génétique et sélection des caractères
2.1.
Les caractères maternels
La sélection
pour accroître la taille de la portée basées
sur le nombre d'ovules pondus pourrait être plus efficace
que la sélection classique sur la taille de portée.
Mais il faudra attendre un plus grand nombre de générations
pour savoir si la méthodes est réellement plus efficace.
|
Deux
contributions s'intéressent à la productivité
numérique au sens large : RUBIO-RUBIO et al.
[23] démontrent la supériorité des femelles
Néo-Zélandaises, pure ou croisées, en prolificité
et poids de portée, dans un protocole visant à comparer
les performances des souches Néo-Zélandaises, Californiennes,
Chinchilla et Criollo. BRUN et BASELGA [4]
ont comparé les performances de la souche 2666, lignée
composite issue du croisement entre la souche 2066 de l'INRA et la
souche " V " de Valence, aux performances de la souche "
V " pure. Un effet d'hétérosis significatif a été
mis en évidence pour le taux de gestation et la prolificité
jusqu'en 4e génération.
Plus nombreuses
sont les contributions s'attachant à étudier les composantes
élémentaires de la prolificité, taux d'ovulation
et survie embryonnaire :
Une expérience de sélection sur le taux d'ovulation,
mesuré par laparoscopie 12 jours après une insémination
artificielle (IA) a été mise en œuvre par IBANEZ
et al. [11]. Après
deux générations de sélection, l'estimation
de la réponse à la sélection est de 0,97 ovules,
0,79 embryons implantés et 0,32 lapereaux nés totaux
par génération. Les précisions de ces estimations
sont faibles et il faut considérer ces résultats comme
préliminaires. En revanche, PEIRO et al. [18]
ne trouvent pas de différences significatives de survie embryonnaire
entre les deux lignées de l'expérience de sélection
divergente sur la capacité embryonnaire conduite pendant
10 générations à l'université de Valence.
Des analyses de ségrégation, visant à mettre
en évidence une transmission héréditaire de
type monogénique, réalisées sur les animaux
de cette expérience, ont cependant suggéré
l'existence d'un gène majeur agissant sur la survie embryonnaire,
comme le mentionnent GARREAU et al. [2]
dans leur rapport de synthèse. Les chercheurs de l'INRA ont
également mis en évidence l'influence du gène
de la caséine Kappa sur la prolificité des femelles
et plus particulièrement sur la survie embryonnaire. La carte
génétique du lapin, en cours d'élaboration
[5], sera un outil privilégié
pour localiser et identifier ces gènes majeurs.
|
| |
Les composantes
mâles et femelles de la fertilité ont fait l'objet
de deux communications :
GARCIA et al. [8] ont estimés
les paramètres génétiques (effets génétiques
directs et maternels, hétérosis) des caractéristiques
de la semence dans les souches mâles espagnoles " C "
et " R " sélectionnées pour la croissance.
Des différences significatives d'effet génétique
direct ont été mis en évidence entre les 2
souches pour les caractères quantitatifs (concentration et
nombre total de spermatozoïdes) et pour les caractères
qualitatifs (% de spermatozoïdes vivants, % d'anomalies morphologiques).
Les effets d'hétérosis n'étaient significatifs
que pour la gouttelette cytoplasmique proximale. Il n'a pas été
montré de supériorité des mâles croisés
par rapport aux mâles de souche pure.
PILES et al. [19] ont abordé
l'étude de la fertilité à l'aide d'une méthodologie
innovante et intéressante : La fertilité a été
analysée en tant que caractère binaire (0 pour les
femelles inséminées non gestantes ; 1 pour les femelles
gestantes) à l'aide d'un modèle introduisant une valeur
génétique pour la femelle et une valeur génétique
pour le mâle. Les héritabilités estimées
(0,06 pour la composante femelle et 0,01 pour la composante mâle)
suggèrent la possibilité d'une sélection pour
la fertilité mais avec une espérance de progrès
génétique faible.
|
|
Homogénéité
du poids
de naissance
|
Lignées |
| + |
- |
| Nés totaux |
8,40 |
9,60 |
| Nés vivants |
7,96 |
8,84 |
| Mortinatalité% |
5,24 |
7,92 |
| Poids naiss. g |
63,6 |
65,2 |
| Coef.Variat. % |
10,9% |
12,4% |
| Sevrés/M.Bas |
7,31 |
7,51 |
| Mort. N-sev.% |
8,2 |
15,0 |
Caractéristiques des portées de
2e génération sélectionnées pour une
forte ou une faible homogénéité du poids à
la naissance [9] (Garreau et al.,
2004)
|
La
sélection pour l'homogénéité des poids
de lapereaux à la naissance fait également appel
à une méthodologie très innovante [9]
: Un modèle introduisant une valeur génétique
pour la moyenne des poids et une valeur génétique pour
la variabilité des poids a permis de créer à
l'INRA une lignée à poids de lapereaux homogènes
et une lignée à poids de lapereaux hétérogènes.
Ainsi le coefficient de variation du poids à la naissance est
réduit de 1,5 points (P<0,05) dans la lignée homogène,
sans variation significative du poids des lapereaux à la naissance
ou au sevrage. La mortinatalité et la mortalité naissance-sevrage
sont significativement plus faibles dans la lignée homogène.
Ces résultats très prometteurs conduiront à une
application chez les professionnels de la sélection dans un
avenir proche.
La méthodologie
de l'analyse de survie se généralise pour analyser
la durée de la carrière productive des femelles. Un
protocole de sélection divergente sur la longévité
des femelles est en cours à l'INRA et les estimations d'héritabilité
réalisées dans les souches françaises et espagnoles
permettent d'envisager la sélection de ce caractère
dans un avenir proche [8]. La valeur
nulle de la corrélation génétique estimée
par SANCHEZ et al. [25] entre
la durée de survie productive des femelles et leur prolificité
confirme l'indépendance génétique de ces deux
caractères.
|
|
2.2.
Les caractères de croissance et de composition corporelle
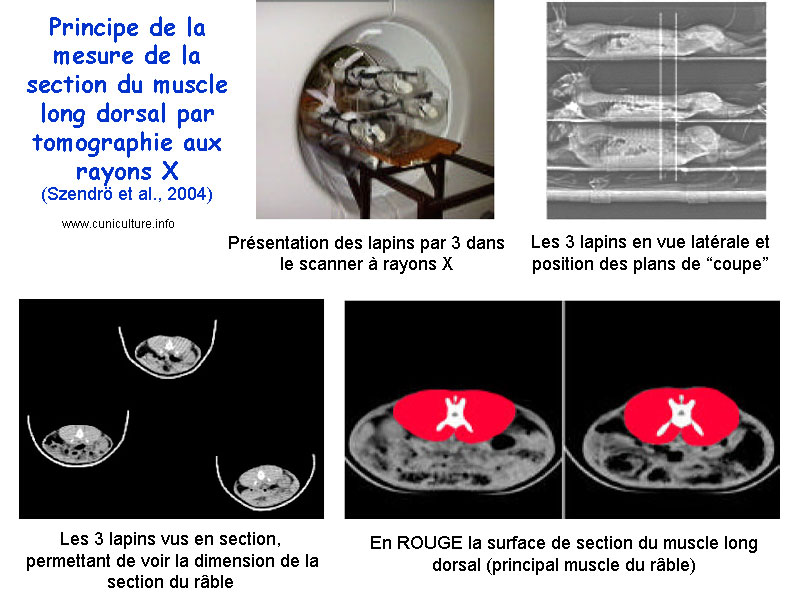
La méthode
de tomographie aux rayons X utilisée pour sélectionner
sur la section du muscle long dorsal (râble)
|
ORENGO et
al. [17]
ont comparé les performances de croissance de 25 types
génétiques issus de croisement réciproques
entre 5 souches espagnoles (3 souches femelles et 2 souches mâles).
Globalement les types génétiques issus des lignées
mâles, sélectionnées pour la croissance, présentaient
un poids plus élevé, une vitesse de croissance plus
rapide, une quantité d'aliment ingérée plus
forte et un indice de consommation plus faible. SANCHEZ et al.
[26] ne trouvent toutefois pas de
réponse corrélée de l'indice de consommation
à une sélection pour le gain de poids quotidien. Ce
résultat, ainsi que la très faible réponse
directe estimée par l'utilisation d'un témoin congelé,
peut s'expliquer par l'utilisation d'un aliment spécifique
pour lutter contre l'entérocolite.
SZENDRÖ
et al. [27] ont mis en œuvre
une sélection combinée sur la vitesse de croissance
et la surface du muscle long dorsal, mesurée in vivo par
tomographie aux rayons X. Les héritabilités et corrélations
génétiques de la mesure de tomographie et du rendement
de carcasse sont élevées (0,41 - 0,79 et 0,45 respectivement)
et laissent envisager une sélection efficace de la conformation
des carcasses. Cette efficacité est confirmée par
les évolutions génétiques du caractère
estimées par la méthode du BLUP dans la population
sélectionnée.
Une autre
expérience de sélection réalisée à
l'INRA concerne l'efficacité alimentaire [13].
Une régression de la consommation alimentaire a été
réalisée sur le gain de poids, le poids métabolique
moyen et une valeur de la teneur en lipides donnée par la
Tobec (Total Body Electrical Conductivity) dans une lignée
mâle. Les animaux ont été sélectionnés
de façon divergente pour la consommation alimentaire résiduelle
calculée par cette régression, considérée
comme un critère de l'efficacité alimentaire. La différence
de consommation résiduelle entre les 2 lignées n'était
pas significative après une génération de sélection
mais la sélection n'a été réalisée
que sur la voie mâle et le nombre d'animaux mesurés
était assez faible. L'héritabilité de ce caractère
reste élevée (0,45 estimée et 0,25 réalisée)
et favorablement corrélée à la croissance et
à l'indice de consommation. Ces travaux devront être
poursuivis mais l'estimation du gras corporel devra être réalisé
différemment car la mesure du Tobec s'est révélée
trop imprécise chez les lapins de 65 jours.
|
|
2.3.
La résistance aux maladies
Il
semble bien exister un variabilité génétique
de la résistance des lapins aux entéropathies
|
Cette thématique
émergente est certainement amenée à se développer
dans les prochaines années en raison de la forte diffusion
des pathologies et parce qu'elle peut constituer une alternative
à l'utilisation massive d'antibiotique.
La variabilité génétique de la résistance
à 3 types d'entéropathie chez le lapin en croissance
a été estimée par de ROCHAMBEAU et al.
[21] : Les mortalités et
morbidités de 2 bandes de lapereaux issus de 48 pères
ont été analysées après 3 challenges
: inoculation de coccidie, régime alimentaire très
pauvre en fibre (ndlr: environ 7% de cellulose
brute et 19% de NDF) et administration de l'inoculum Tech-2
de l'INRA (permettant de reproduire l'Entéropathie
Épizootique du Lapin - EEL). L'effet du père
était significatif pour la mortalité et la morbidité
dans les 2 premiers traitements mais pour la morbidité seulement
dans le 3e cas. Le classement des pères était similaire
pour la mortalité et la morbidité entre les 2 premiers
challenges mais seulement pour la morbidité entre le premier
et le troisième. Ces résultats semblent démontrer
des mécanismes de résistance différents entre
d'une part l'inoculation de coccidie et le régime pauvre
en fibre, d'autre part l'inoculation de l'inoculum Tech-2, au moins
pour la mortalité. Malgré tout, il semble exister
une variabilité génétique de la résistance
aux entéropathies.
|
| Il
semble qu'il existe aussi une résistance génétique
des lapins à la pasteurellose. Toutefois l'imprécision
de l'évaluation des symptômes rend la sélection
de lignes résistantes encore impossible au plan pratique. |
L' étude
menée par EADY et al. [7]
avait pour but de vérifier si les observations relatives
à l'état de santé général des
lapins, réalisées lors du tri en fin d'engraissement,
pouvaient être utilisées pour améliorer la
résistance génétique aux maladies bactériennes,
et en particulier à la pasteurellose. L'héritabilité
de l'incidence de cette maladie, mesurée sur des lapins en
croissance issus de 2 souches mâles du sélectionneur
Grimaud, variait de 0,04 à 0,15 selon la méthode utilisé,
suggérant la possibilité d'une sélection pour
la résistance. De nouveaux développements méthodologiques
et un enregistrement plus précis des symptômes devront
être réalisés avant d'utiliser ces observations
pour l'amélioration génétique de la résistance
à la pasteurellose
|
|
2.4
Adaptation aux climats chauds

|
HASSAN [10]
a estimé les paramètres génétiques des
caractères de croissance de lapins Néo-Zélandais
élevés en Égypte. Les héritabilités
présentées semblent surestimées, probablement
en raison d'un problème d'estimation de l'effet d'environnement
commun dont la valeur est anormalement faible.
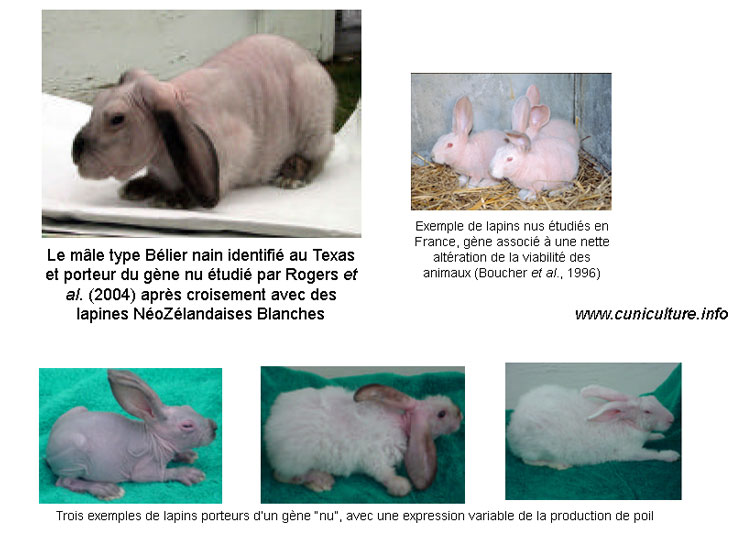
ROGERS et
al. [22] ont comparé
dans les conditions subtropicales du Sud Texas, les performances
de croissance de lapins sans pelage, porteurs d'un gène nu
isolé aux USA, à leurs homologues à fourrure
. Les lapins nus avaient un poids final plus élevé,
une meilleure croissance (gmq de 27,3 vs 24 g/jour - P<0,001)
et une consommation alimentaire plus forte mais un indice de consommation
plus élevé (4,35 vs 3,34 sur
6 semaines à partir de l'âge de 30-35 j). De
plus la température rectale et la fréquence respiratoire
étaient classiquement plus faibles chez les lapins nus, démontrant
une meilleure tolérance de ces animaux à de fortes
températures (ndlr: mais aussi une
moindre tolérance au froid).
|
|
3.
Mise en œuvre des biotechnologies
La
carte génétique du lapin est bien avancée.
|
La
contribution la plus intéressante de ce volet pour la filière
du lapin de chair est sans aucun doute l'élaboration de
la carte génétique et cytogénétique du
lapin :
En 2001, 3 laboratoires de l'INRA ont lancé le projet de la
cartographie du génome du lapin [5].
Le but de ce projet était de produire des marqueurs génétiques
répartis uniformément sur l'ensemble du génome
(carte génétique) et de localiser ces marqueurs sur
les chromosomes (carte cytogénétique). Nous disposons
aujourd'hui de 178 microsatellites localisés, répartis
sur tous les chromosomes sauf le 21. Il n'est pas prévu de
rechercher de nouveaux microsatellites dans l'immédiat mais
nous disposons d'une centaine de microsatellites supplémentaires,
exploitables avec des amorces fluorescentes, et le séquençage
du génome du lapin, qui sera vraisemblablement terminé
à la fin de l'année 2005, permettra également
d'enrichir la carte. A terme, cette carte permettra d'identifier des
gènes d'intérèt et de les introgresser dans les
populations ou de réaliser de la sélection assistée
par gène.
Deux contributions
concernaient la transgenèse chez le lapin dans le cadre de
recherches biomédicales [6,20].
L'une s'intéressait à la stabilité et au taux
de transmission du transgène introduit chez le lapin. Sur
3 générations incluant des croisements avec des sujets
non porteurs du transgène, la stabilité de la transmission
de ce transgène aux descendants a été confirmée.
L'autre contribution s'intéressait à la croissance
et la production laitière de lapins transgéniques.
Les animaux transgéniques se sont avérés un
peu plus lourds (1-30 jours) que les autres, et leur production
laitière un peu plus élevées que celles des
sujets non-transgéneniques, mais les différences n'étaient
pas significatives..
Enfin, le séquençage
des gènes du complexe majeur d'histocompatibilité
[14] a permis de montrer un polymorphisme
élevé susceptible d'être mis à profit
pour la sélection et notamment pour la résistance
aux maladies.
|
|
Conclusion
|
La
prolificité demeure un objectif primordial pour l'amélioration
des souches femelles mais la sélection s'attache aujourd'hui
à améliorer la longévité des femelles,
le poids et la survie des lapereaux, la fertilité et la résistance
aux maladies. L'amélioration de la croissance reste une voie
privilégiée pour les souches mâles mais les recherches
s'orientent également vers l'amélioration de la qualité
des carcasses et de la semence, de l'efficacité alimentaire
et de la résistance à l'entérocolite. La carte
génétique du lapin, actuellement en cours de développement,
sera l'outil de choix pour une meilleure connaissance de la variabilité
génétique de ces caractères. L'organisation de
la sélection vise à améliorer l'efficacité
des schémas de sélection et la diffusion du progrès
génétique vers les élevages afin de mieux répondre
aux attentes de la filière. |
| |
|
|
|