| |
G.
COUREAUD1,
D. MONTIGNY1,
A.S. MONCOMBLE1,
B. PATRIS1,
G. PERRIER2, B. SCHAAL1,
2005. Apprentissages olfactifs du lapereau sous la mère.
11èmes
Journées de la Recherche cunicole, 29-30 nov. 2005 Paris, ITAVI
édit., 33-36.
1 Centre
Européen des Sciences du Goût, Equipe d’éthologie,
UMR 5170, CNRS, 21000 Dijon, France
2 Etablissement National d'Enseignement Supérieur Agronomique,
21000 Dijon, France. |
Résumé - Les odeurs jouent un rôle clé
dans l’adaptation du lapereau nouveau-né, en influant
notamment sur son comportement alimentaire. Des mécanismes
prédisposés et des mécanismes d’apprentissage
sous-tendent l'orientation du lapereau vers la mère et le
succès des tétées. Ces mécanismes fonctionnent
en interaction, mais peuvent être dissociés expérimentalement.
La signalisation mammaire repose sur un signal phéromonal,
actif indépendamment de tout apprentissage. Cette phéromone
mammaire agit non seulement comme un signal de déclenchement
du comportement de recherche de la mamelle, mais aussi comme un
agent facilitateur des apprentissages olfactifs. Par cette fonction,
la phéromone mammaire est en mesure de contribuer à
l’amélioration chez le lapereau des performances de
tétée et de reconnaissance des congénères,
et de le préparer aux transitions alimentaires et sociales
à venir.
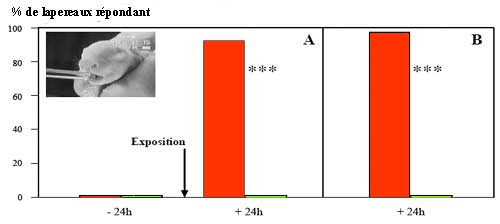
| Exemple
de résultat de test : Proportions de lapereaux de
2 jours exprimant le comportement de recherche de la mamelle
dans un test d’activation orale les exposant à différents
odorants sur une baguette de verre (cf. photo). Cas A:
les stimulations sont constituées d'un support neutre
d’éthyl-acéto-acétate (EAA; barres
rouges) ou de furanéol (barres vertes). Le test est effectué
24h avant (pour vérifier la neutralité des supports)
puis après exposition des lapereaux au mélange
odorant phéromone mammaire + EAA : les lapereaux sont
clairement attirés par le mélange phéromone
+ EAA et non par le témoin. Cas B :le test est
effectué 24h après exposition des lapereaux à
l’EAA seul, mais au préalable, celui-ci a été
badigeonné sur le ventre de la mère avant la tétée.
Les lapereaux sont attirés par l'EEA dont ils avaient
senti l'odeur sur le ventre de leur mère et non par le
témoin furanéol (n=40 lapereaux/essai). |
|
|
|
|
|
| |
|
|
| |
|
R.
NOWAK, J. SERRA, 2005. Appren-tissages des caractéristiques
olfactives du nid chez le lapereau nouveau-né. 11èmes
Journées de la Recherche cunicole, 29-30 nov. 2005 Paris,
ITAVI édit., 37-40.
UMR 6175,
PRC, INRA CEntre de Tours, 37380 Nouzilly, France.
|
Résumé
:
De résultats suggérant une discrimination positive
précoce du nid familial par le lapereau, a émergé
l’hypothèse d’un apprentissage très précoce
des indices olfactifs de cette niche écologique particulière.
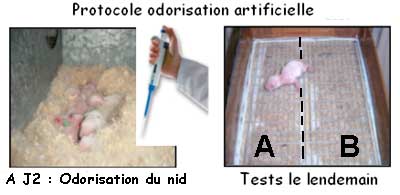
Une expérience consistant à odoriser le nid avec du
thym ou de la cannelle le deuxième jour après la naissance
indique que le nouveau-né est attiré vers l'odeur
placée dans son nid lors du test effectué le lendemain.
Ainsi ce dernier est capable d’enregistrer, très rapidement
et très tôt dans sa vie, de nouvelles caractéristiques
odorantes relatives à son nid. En outre, diverses comparaisons
mettant en jeu des nids odorisés ou non, familial ou "étranger",
le contenu de la boite à nid familiale prélevé
avant ou après odorisation, ont permis de démontrer
l’apparition d’une préférence pour les éléments
odorants les plus récents lorsque ceux-ci étaient
comparés à ceux présents dès la naissance.
Pour chaque test comparant deux sources potentielles d'odeurs, 20
lapereaux issus de 10 portées différentes ont été
utilisés
 |
Lors du test,
un lapereau est déposé au milieu d'une grille
au dessous de laquelle a été placée en
A et en B soit de la litière + du poil + etc. ... provenant
de différents nids, soit des tampons porteurs des odeurs
artificielles. Les déplacements du lapereau sont suivis
pendant 5 mn et le temps total passé au dessus de chaque
moitié est enregistré. Un lapereau n'est "utilisé"
qu'une seule fois. |
|
|
|
| |
|
|
| |
|
J. SERRA, R. NOWAK, 2005. Développement
d’une préférence pour l’environnement postnatal
chez le lapereau nouveau-né. 11èmes
Journées de la Recherche cunicole, 29-30 nov.
2005 Paris, ITAVI édit., 41-44.
Equipe
comportement, UMR 6175, PRC, INRA Centre de Tours 37380 Nouzilly,
France
|
Résumé
:
L’objectif de ce travail était d’évaluer
les capacités d’orientation olfactives du lapereau envers
son environnement postnatal. Des lapereaux ont été
placés dans des situations de choix au premier (J1) et au
septième jour (J7) après la naissance. Dès
le premier jour, le nouveau-né est attiré par sa mère
et par toute autre lapine allaitante, de même que par sa fratrie
ou tout autre lapereau étranger du même âge.
Il s’oriente aussi préférentiellement vers son
nid ou un nid étranger lorsque ceux-ci sont mis en comparaison
avec un compartiment vide. Dans un test de choix présentant
deux stimuli de manière concomitante, il y a reconnaissance
du nid familier par rapport au nid étranger à J1 et
à J7. En revanche, aucune préférence n’a
pu être montrée pour la mère par rapport à
une lapine étrangère au même stade physiologique,
ou pour la fratrie par rapport à des lapereaux étrangers
du même âge.

Dispositif à 2 compartiments, ouvert et prêt à
recevoir des lapereaux ou le contenu de nids pour les tests |

Réalisation par un lapereau du test de choix entre 2
nids (son propre nid et un nid étranger, placés
chacun dans un compartiment) |
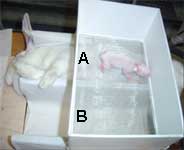
Réalisation par un lapereau du test de choix entre rien
(B) et une lapine (A) présentée couchés
sur le dos |
|
|
|
| |
|
|
| |
|
J.
ORENGO*, T. GIDENNE, 2005. Comportement alimentaire et caecotrophie
chez le lapereau avant sevrage. 11èmes Journées
de la Recherche cunicole, 29-30 nov. 2005 Paris, ITAVI édit.,
45-48 .
INRA, Station
de Recherches Cunicoles, BP 52627, 31326 Castanet-Tolosan Cedex,
France
*Adresse actuelle: Departamento de Producción Animal,
Facultad de Veterinaria, Universidad de Murcia, Espagne
|
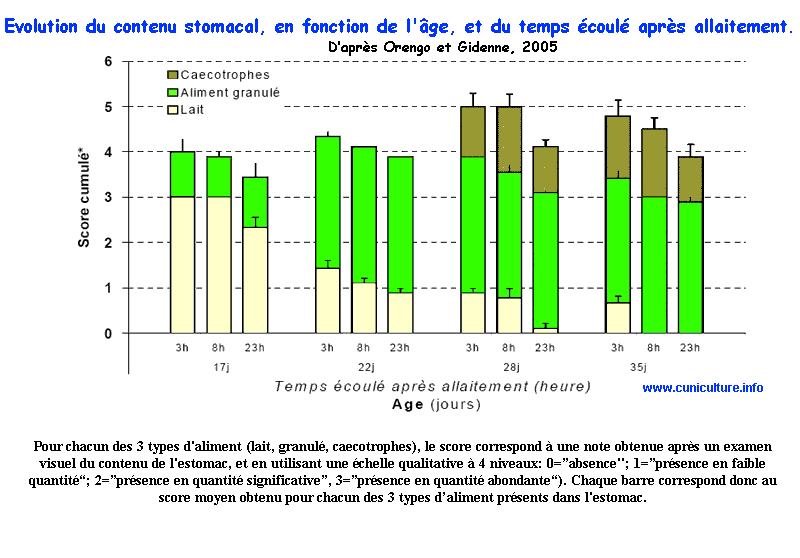
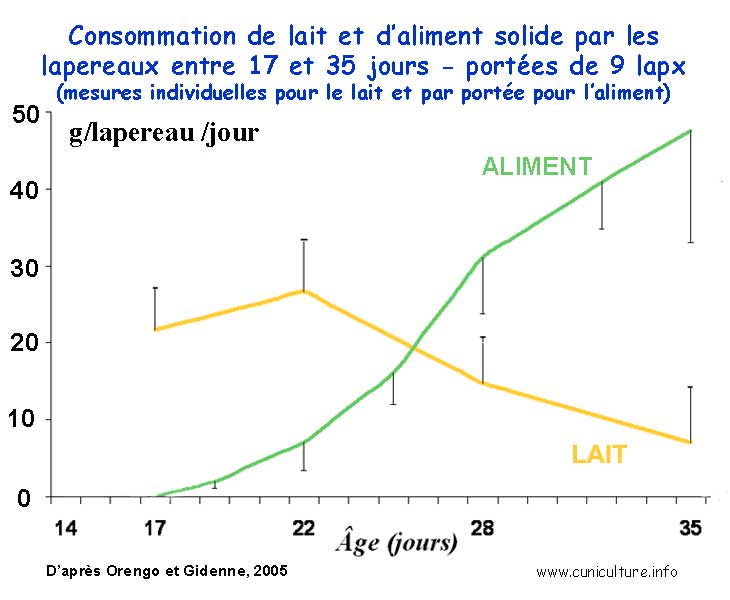
Résumé
- L'étude implique 21 portées de femelles primipares
(NZW x Cal), égalisées à 9 lapereaux à
la naissance. Chaque portée est logée avec sa mère
dans une cage permettant une alimentation séparée.
L'ingestion de lait et d'aliment, ainsi que le contenu de l'estomac
ont été mesuré à différents âge
(17-22-28-35 jours) et en fonction du temps écoulé
après allaitement (3-8-23h). La nature du contenu stomacal
a été chaque fois notée pour la présence
de lait, d'aliment et de caecotrophes selon un échelle allant
de 0 (absence) à 4 (présence importante). Les lapereaux
consomment quasi-exclusivement du lait jusqu'à l'âge
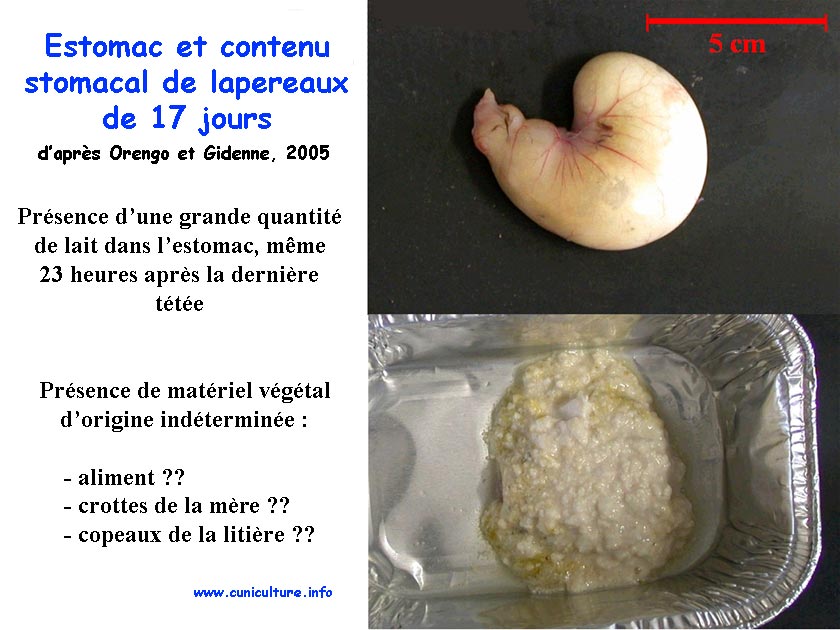
de 17 jours. Dans l'estomac, 23h après allaitement, on observe
une quantité importante de lait à 17 et 22 jours (score=2,3
et 0,9 resp. à 17 et 22j) et pratiquement plus à 28
ou 35 j. Dès 17 jours d'âge une présence peu
importante d'aliment est observée dans l'estomac (score=
1,0), puis celle ci s'accroît fortement dès 22 jours
d'âge. A partir de 17j, le lait coagulé est surtout
présent dans le fundus, et en périphérie de
l'estomac (proche de la muqueuse), tandis que l'aliment est réparti
de manière plus homogène (tant qu'il y a absence de
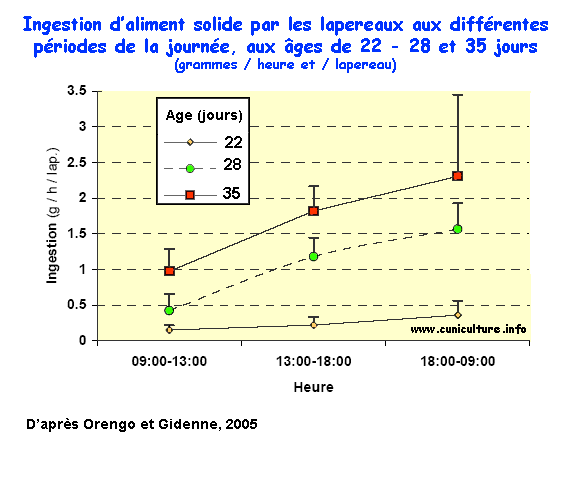
caecotrophes). Dès 22 j, l'ingestion d'aliment granulé
atteint 7,2 g/j/lap., puis passe à 31 et 48 g/j respectivement
à 28 et 35 jours, mais elle reste très variable d'une
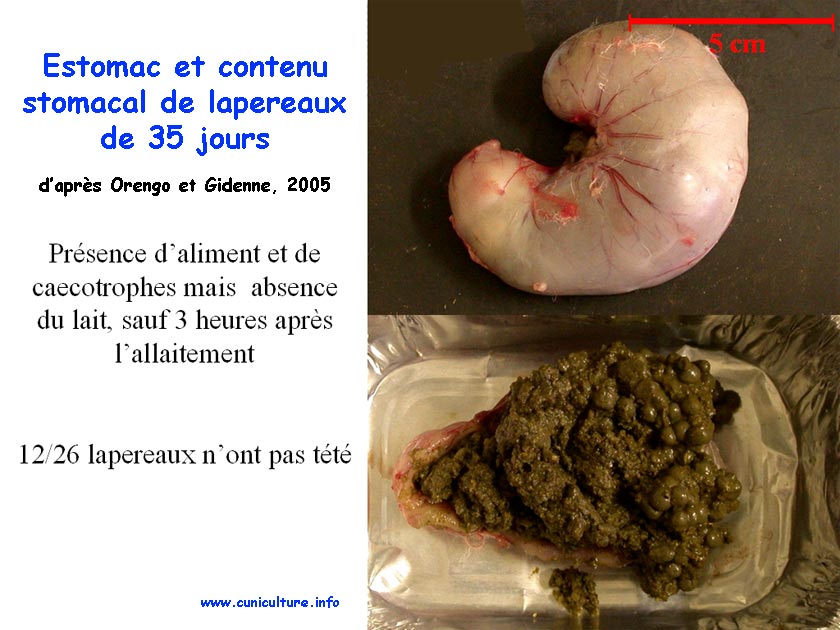
portée à l'autre (CV=30% à 35 j). Entre 22
et 35 jours d'âge, l'ingestion d'aliment s'accroît avec
le délai post-allaitement, et 73 à 76% de l'ingestion
d'aliment est effectuée entre 18:00 et 09:00h, quel que soit
l'âge. Ainsi, le comportement alimentaire nocturne s'installe
dès que le lapereau débute l'ingestion d'aliment solide.
Dès 28 j, des caecotrophes sont trouvés dans le fundus
et représentent 25 à 30% du contenu stomacal. Enfin,
tandis que le poids du contenu frais de l'estomac double entre 17
et 35 jours, la teneur en matière sèche du contenu
stomacal décroit de 31,1% à 17 jours à 19,1%
à 35 jours. Elle est également plus élevée
3 et 8 heures après l'allaitement (26,8 et 27,9%) que 23
heures après (21,5%).
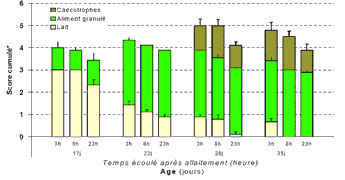 Evolution
du contenu stomacal, en fonction de l'âge, et du temps
écoulé après allaitement
Evolution
du contenu stomacal, en fonction de l'âge, et du temps
écoulé après allaitement |
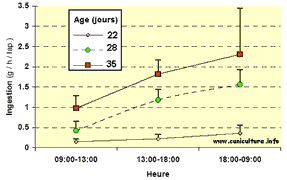 Ingestion
d'aliment solide par les lapereaux aux différentes
périodes de la journée, aux âges de 22
- 28 et 35 jours (g/heure &/lapereau)
Ingestion
d'aliment solide par les lapereaux aux différentes
périodes de la journée, aux âges de 22
- 28 et 35 jours (g/heure &/lapereau) |
|
|
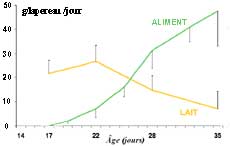
Ingestion de
lait et d'aliment solide par les lapereaux entre 17 et 35 jours
(portées de 9)
.
|
|
|
| |

Estomac et contenu
stomacal de lapereaux de 17 jours |
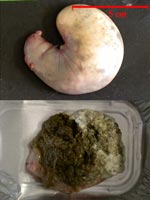
Estomac et contenu
stomacal de lapereaux de 22 jours |
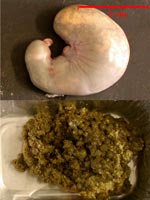
Estomac et contenu
stomacal de lapereaux de 28 jours |
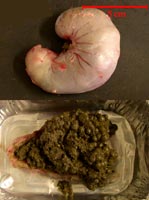
Estomac et contenu
stomacal de lapereaux de 35 jours |
|
|
| |
|
|
| |
|
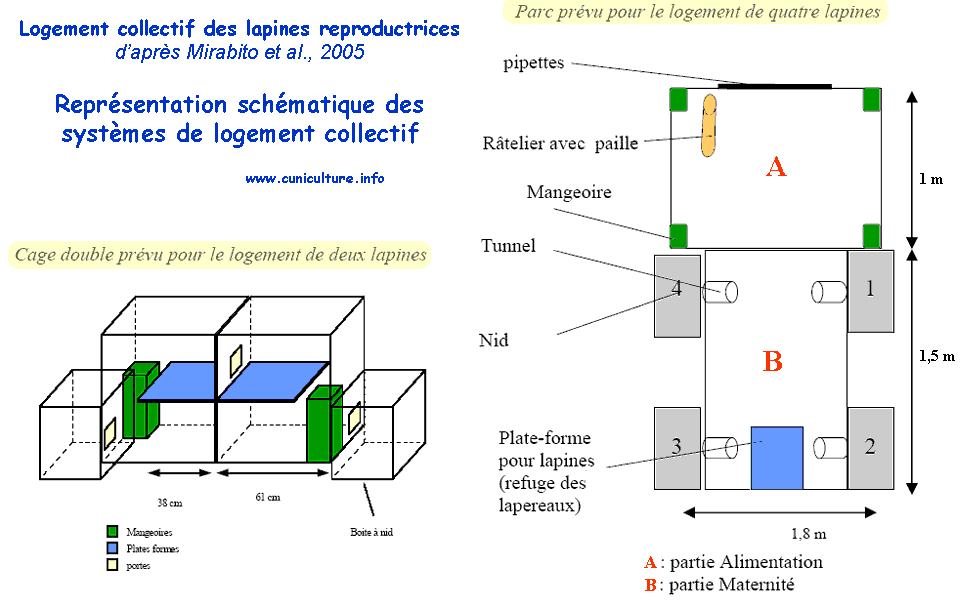
L.
MIRABITO, C. BOQUIER, 2005. Développement d’un système
d’alimentation séparée des mères et des
jeunes avant sevrage. 11èmes Journées de la Recherche
cunicole, 29-30 nov. 2005 Paris, ITAVI édit., 49-52 .
ITAVI 28, Rue
du Rocher 75008 Paris, France.
Travail réalisé
avec la collaboration des Ets Arrivé Bellané, Cavac,
Chabeauti, Inzo et Trouw Nutrition France
|
Résumé.
L’évolution du mode de logement des lapines qui pourrait
être induite par de futures réglementations sur le
bien-être des lapins, notamment une augmentation de la hauteur
des cages, a permis la mise au point d’un nouveau système
d’alimentation séparée des mères et des
jeunes pendant la lactation à la station expérimentale
de l’ITAVI. Dans la présente étude, nous rapportons
les résultats obtenus dans un site où 30 femelles
et leur portées ont été logées dans
des cages expérimentales et leurs performances comparées
au cours de 4 essais successifs avec un échantillon de femelles
« témoin ». Les cages expérimentales se
caractérisaient par une hauteur de 60 cm avec présence
d'une plate-forme grillagée d'une longueur de 30 cm située
contre la face arrière de la cage à 32 cm de haut.
La longueur des cages était de 95 cm (64 cm sans l’espace
nid) et la largeur de 42 cm. La mangeoire des lapines était
positionnée de telle façon que le bord inférieur
de la partie accessible soit situé à 32-34 cm de haut.
Les lapines accédaient à l'aliment en se redressant
sur leurs pattes postérieures (photo ci-dessous). La mangeoire
des lapereaux était de conception classique (12 cm de profondeur
et 16 cm de largeur avec deux postes). L'accès des lapines
à l'aliment des lapereaux n'était pas possible en
raison de la présence d'une tige métallique horizontale
qui limitait la hauteur de l'accès à 5 cm au niveau
de cette mangeoire (photo ci-dessous).
Il ressort
de ce travail que, jusqu'à 18-19 jours d’âge des
lapereaux, le logement expérimental n’a aucun effet
sur les performances zootechniques. Au delà, une réduction
du poids au sevrage des lapereaux a été observée
dans un essai sur quatre. Celle-ci nous semble devoir être
mise en relation avec une réduction de la consommation des
lapereaux dans le système expérimental. Nous émettons
l’hypothèse que, dans ce système d’alimentation,
les performances de croissance des lapereaux, au delà de
18-19 jours, sont dépendantes de leur capacité à
consommer précocement l’aliment granulé. Dans
ce cadre, une mise en place de la mangeoire des lapereaux dès
14 jours pourrait constituer un des éléments essentiels
à sa réussite.
|
|
|
| |
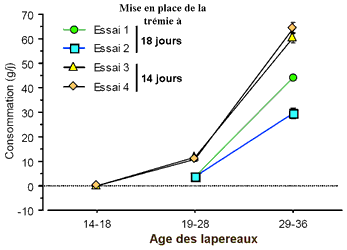
Consommation alimentaire
des lapereaux dans les 4 essais successifs |

La lapine dressée
sur ses pattes pour accéder à sa mangeoire |

La mangeoire des
lapereaux au premier plan. Celle de la mère en hauteur à
l'arrière |
|
|
| |
|
|
| |
L.
MIRABITO, P. GALLIOT, C. SOUCHET, F. DUMONT, F. THOMERET, 2005. Logement
collectif des lapines repro-ductrices : Conséquences zootechniques.
11èmes
Journées de la Recherche cunicole, 29-30 nov. 2005 Paris, ITAVI
édit., 53-56 .
ITAVI 28, Rue du
Rocher 75008 Paris, France. |
Résumé. Les performances zootechniques des
lapines logées en cage individuelle conventionnelle (n=27),
par paires en cages rehaussées avec plate-forme (n=27 groupes)
et en parc collectif de 4 femelles (n=19 groupes) ont été
suivis au cours des quatre premiers cycles de reproduction. Globalement,
le mode de logement n’a pas eu d’incidence sur la fertilité,
la prolificité des lapines et la croissance des lapereaux.
Il a seulement été mentionné une meilleure
fertilité des lapines logées en cage individuelles
que da,s les deux systèmes collectifs (80% vs 51% P=0,04).
En revanche, en parc, la mortalité des lapereaux entre 3
et 33 jours était significativement plus élevée
que celle observée en cage individuelle au cours de deux
cycles. [NDLR : un calcul complémentaire
à partir des données des auteurs montre que pour les
4 cycles pris dans leur ensemble, la mortalité globale des
lapereaux entre 3 et 33 jours a été très significativement
supérieure (test de khi² : P<0,001) en parcs (17,6%)
par rapport à celle observée dans les cages doubles
(10,6%) ou individuelles (8,7%), ces 2 derniers modes d'élevage
ne se distinguant pas.] Ce résultat semble principalement
s’expliquer par la fréquence élevée, observée
en parc, des mises bas multiples dans le même nid (37,5 %
des mises bas). Enfin, cet essai s’est caractérisé
par la fréquence élevée des blessures/abcès
observés chez les femelles (30 % des causes de réforme)
et liées principalement à un élevage collectif
des futures reproductrices.
|
|
|
| |
|
|
| |
|
|
| |
L.
MIRABITO, F. DUMONT, P. GALLIOT, C. SOUCHET, 2005. Logement collectif
des lapines reproductrices Conséquences sur le comportement
. 11èmes
Journées de la Recherche cunicole, 29-30 nov. 2005 Paris, ITAVI
édit., 57-60 .
ITAVI 28, Rue du
Rocher 75008 Paris, France |
Résumé.
Le comportement des lapines a été évalué
au cours du premier cycle de production dans trois mode de logement
: individuel en cage conventionnelle (n=18), par paire en cages
rehaussées avec plate-forme (n=9) et par groupe de quatre
en parc (n=9). [cf. communication précédente].
L'identification des activités a été réalisée
chaque semaine lors de 7 séquences d'observation de 20 minutes
réparties sur la totalité du cycle de 24h. Globalement,
le temps passé en cohésion par les animaux dans les
deux logements collectifs est apparu très différent
: 0,8 % dans les logements par paire et 29,9 % dans les groupes
de quatre lapines ce qui se traduisait aussi par une fréquence
des interactions sociales, quelle que soit leur nature, significativement
supérieure en parc. Dans les logements collectifs, malgré
l’augmentation de la hauteur et de la surface disponible, les
postures étaient peu modifiées par rapport à
celles relevées en cage individuelle : possibilité
d’adopter une position « redressée » (0,5%
du budget-temps) et temps passé à se déplacer
significativement supérieur dans les parcs par rapport aux
deux autres modes de logement (2,7 % vs 1,2 et 0,6 % - P<0,002).
Enfin, en terme d’activités, seules quelques différences
ponctuelles ont été relevées et le logement
en parc n’est pas apparu de nature à réduire
la fréquence des tentatives d'allaitement ou de mordillage
des cages par rapport au logement individuel. En conclusion, le
logement collectif des lapines reproductrices n’est pas apparu
au cours de cette étude de nature à modifier sensiblement
le comportement des lapines. S’il permet l’expression
d’un comportement plus diversifié et d’interactions
sociales notamment en parc, ces bénéfices sont cependant
contre balancés par l’impact négatif enregistré
par ailleurs sur la viabilité des lapereaux en parcs [cf.
communication précédente] ou des femelles fortement
dominées dans les logements par paire.
|
|
|
| |
|
|
| |
L. MIRABITO, P. GALLIOT,
C. SOUCHET, 2005. Effet de la surface disponible et de l'aménagement
des cages sur les perfor-mances zootechniques et le comportement des
lapines. 11èmes
Journées de la Recherche cunicole, 29-30 nov. 2005 Paris, ITAVI
édit., 61-64 .
ITAVI 28, Rue du
Rocher 75008 Paris, France |
Résumé.
Au cours de cette étude, nous avons évalué l’impact
de la surface disponible (3420, 4508, 5855 cm²) combinée
ou non à l’aménagement des cages (plate-forme ou
tunnel) sur les performances zootechniques et les postures adoptées
par les lapines (10 ou 20 cages étudiées par type).
Le mode de logement n’a induit aucune différence sur les
performances zootechniques mesurées au cours de cinq cycles
de production. De même, durant la phase diurne, des variations
minimes des postures ont été observées en relation
avec la surface disponible alors que la présence d’une
plateforme induisait principalement une substitution entre les différentes
postures « couché » adoptées par les animaux.
En conclusion, cet essai semble montrer que, dans la gamme de variation
testée, la surface disponible par animal a peu d’influence
et nous émettons l’hypothèse que les postures «
couché » adoptées par les femelles sont surtout
dépendantes de la forme des cages. |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|
| |
|
|