| |
|
Le
rapport général de la session Physiologie de la digestion
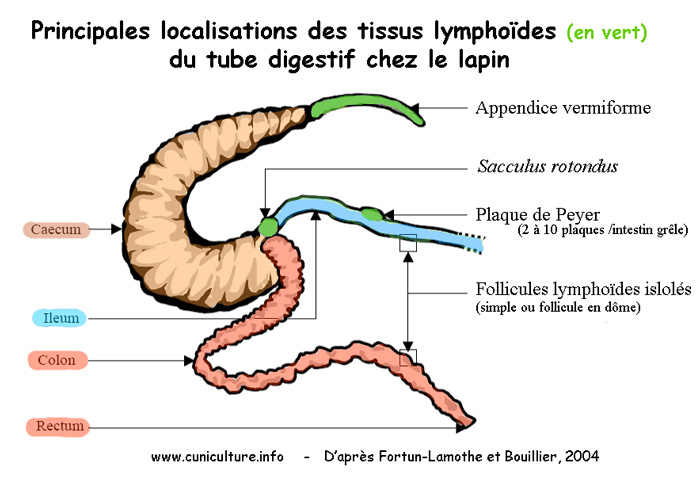
Figure 1
: Localisation des tissus lymphoïdes sur le tube digestif
|
|
Le rapport général
présenté par deux scientifiques INRA/ENV de Toulouse
[1] était consacré
aux interactions entre la flore digestive et l'immunité pariétale
digestive d'une part et aux stratégies envisageables pour
améliorer la santé digestive des jeunes lapereaux
d'autre part. Compte tenu de son ampleur (24 pages de texte et figures
basées sur 140 références), le plus simple
pour le présentation de ce rapport très complet et
détaillé, est de reprendre son résumé
(voir ci-dessous). Les auteurs ont insisté sur le rôle
et la répartition des tissus lymphoïdes et des lymphocytes
qu'ils génèrent. En effet ces cellules sont à
l'origine des défenses immunitaires spécifiques. Elles
sont réparties tout au long de la paroi intestinale et dans
des localisations spécifiques telles que l'appendice cæcal
ou le sacculus rotondus (jonction entre l'intestin grêle et
le cæcum). La localisation macroscopique est schématisée
sur la figure 1
et la localisation microscopique sur la figure
2. Nous en retiendrons surtout que le système de défense
du tube digestif est pratiquement autonome et très complexe,
tous les éléments constituant la paroi du tube digestif,
le chyme alimentaire et la flore digestive interagissant entre eux.
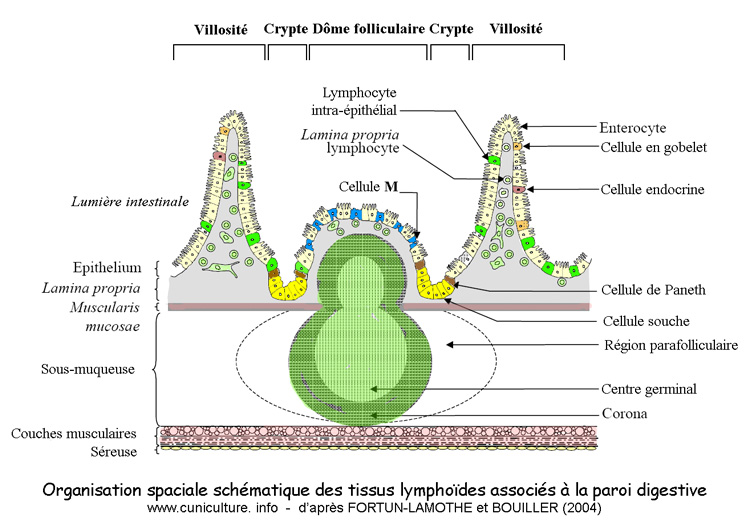
Par exemple les "corps étrangers" présents
dans la lumière intestinale sont reconnus au niveau des cellules
M à la surface des dômes folliculaires ou des plaques
de Peyer et la sécrétion d'IgA effecutée en
réaction pour les neutraliser est assurée dans les
autres parties de la muqueuse digestive (figure
3)
|
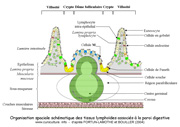
Figure
2: Organisation spaciale schématque des tissus lymphoïdes
associés à la paroi digestive
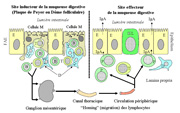
Figure
3 : Organisation fonctionnelle schématique du tissu
lymphoïde associé au tube digestif
|
Résumé
du rapport général : Les troubles digestifs
apparaissent très souvent autour du sevrage et conduisent
à une utilisation importante d'antibiotiques dans l'élevage
du lapin. Dans ce contexte, les éleveurs comme les consommateurs
souhaitent que soient trouvées des stratégies alternatives
permettant d'améliorer la santé des animaux. Cependant
le maintien de la santé du tube digestif est complexe et
dépend de l'équilibre fragile entre la muqueuse (comprenant
l'épithélium qui absorbe les nutriments et le système
immunitaire digestif), la flore commensale et les facteurs d'environnement
qui incluent l'alimentation. Tout d'abord les mécanismes
immunitaires et non-immunitaires de protection contre les agents
pathogènes sont présentés. Sont ensuite présentées
les conditions d'installation et la composition de la flore digestive,
suivies par l'analyse de son rôle dans la santé. Enfin,
plusieurs stratégies sont présentées et discutées;
elle visent à stimuler le système immunitaire digestif
ou à favoriser la flore bénéfique qui élimine
les agents pathogènes digestifs.
Plusieurs nutriments sont impliqués dans le développement
de la réponse immunitaire et pourraient être utilisés
pour favoriser les défenses immunitaires des animaux. Parmi
ceux-ci les acides gras alimentaires (taux et rapport oméga3
/ oméga6) pourraient présenter un certain intérêt
pour les lapins. Le rôle des fibres alimentaires sur la santé
digestive a déjà été démontré
chez le lapin et des rapports étroits entre les apports de
fibres [quantité et qualité] et la microflore cæcale
ont été démontrés. Quelques travaux
ont aussi montré l'influence du taux de fibres de l'aliment
consommé par les lapereaux avant leur sevrage sur leur santé
après ce dernier. C'est pourquoi les besoins alimentaires
des lapereaux allaités, en particuliers leurs besoins en
fibres, font l'objet de nombreux travaux avec comme objectif l'amélioration
de leur santé ultérieure tout en respectant les besoins
de leur mère. Une flore exogène peut aussi être
ajoutée à l'alimentation pour stimuler les défenses
immunitaires digestives des lapins et prévenir le développement
des agents pathogènes. Enfin des vaccins peuvent permettre
la protection de l'hôte contre des agents pathogènes
spécifiques.
|
| |
|
Analyses
de la flore digestive et des causes de sa variation
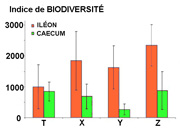
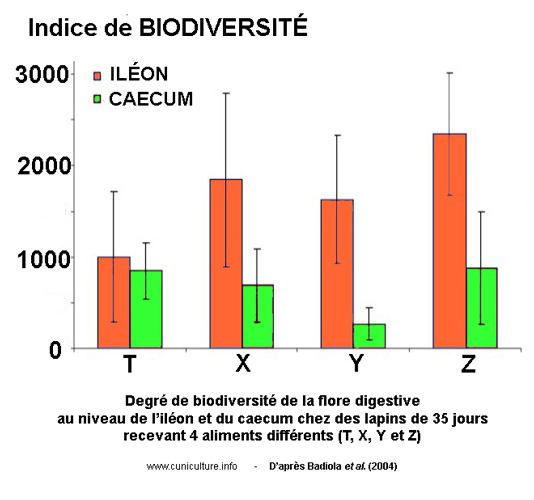
Figure 4 : Indices de biodiversité de la flore iléale
et caecale dans la cas de 4 aliments (T, X, Y, Z)
|
L'apport
des méthodes de biologie moléculaire.
Les méthodes
de biologie moléculaire permettent de séparer l'ADN
des microorganismes et en comparant les différentes séquences
constitutives de cet ADN, elles permettent de déterminer
un index de la biodiversité des microorganismes présents
dans la flore digestive. Le principal avantage est que cette méthodologie
prend en compte la totalité des microorganismes présents
dans la flore et non pas seulement les 20-25% cultivables. La comparaison
des séquences d'ADN obtenues avec celles de banques de données
permet également d'identifier un très grand nombre
de microorganismes.
Par cette méthodologie l'équipe espagnole de Badiola
et al. [12] a confirmé
qu'entre les âges de 15 et 35 jours (avec un point intermédiaire
à 25 j) la nature de la flore évolue, certains microorganismes
apparaissant et d'autres disparaissant, cette évolution étant
indépendante du type d'aliment (4 aliments testés).
L'apport original de ce travail a été de démontrer
que la flore du contenu iléal (fin de l'intestin grêle)
est systématiquement plus diversifiée que celle du
cæcum. Comme le montre la figure 4, l'ampleur de la biodiversité
et l'écart entre iléon terminal et cæcum sont
aussi très dépendants de l'alimentation des lapins.
On doit par exemple remarquer que l'utilisation de l'aliment T a
conduit à une biodiversité proche dans l'iléon
et dans le cæcum, alors que l'utilisation d'un autre aliment
Y a conduit à un écart beaucoup plus important (les
auteurs n'ont donné aucune indication sur la nature des 4
aliment utilisés).
|
| |
La même
équipe, dans un travail sur l'influence du taux de fibres
alimentaires et de leur broyage [24]
a confirmé la plus grande biodiversité du contenu
iléal par rapport au contenu cæcal. En outre, le degré
de similarité entre les flores cæcale et iléale
est de 65%. Un aliment à teneur élevée en fibres
(30% NDF vs 25%) conduit à une réduction de la biodiversité
de 31% sans altération de la digestibilité des fibres,
mais avec une réduction de celle de la matière sèche
et des matières azotées. Un broyage très grossier
(grille de 9 mm vs 1 mm) conduit aussi à une réduction
de cette biodiversité (- 40%), mais cette fois associée
à une réduction de 9% de la digestibilité des
fibres (NDF). Le broyage grossier réduit par exemple dans
la flore iléale les bactéries des genres Escherichia,
Helicobacter ou Klebsiella. L'aliment ayant le taux de
fibres le plus faible réduit de son côté le
nombre de bactéries du genre Bacteroïdes (impliquées
dans la digestion des fibres). Enfin, les auteurs soulignent que
la flore cæcale est nettement moins influencée par
les aliments testés que celle de l'iléon.
|
|
Une séparation
de la mère et de sa portée en dehors des allaitements
contrôlés ne modifique que très transitoirement
la mise en place de la flore digestive des lapereaux entre 0 et
10 jours
|
La
flore digestive analysée par les méthodes classiques
de culture in vitro
Les chercheurs
spécialisés sur le comportement animal (Hudson
et al., 1996) ont montré il y a quelques années
qu'à l'occasion de chaque allaitement la lapine laissait
dans le nid de ses jeunes lapereaux quelques crottes molles. Ces
crottes disparaissent au cours de la journée, probablement
ingérées par les lapereaux.. L'hypothèse a
été émise que leur rôle était
de fournir aux lapereaux une flore digestive similaire à
celle de leur mère. L'équipe hongroise de Kovacs et
al. [6] a voulu vérifier
cette hypothèse. Par des allaitements libres ou contrôlés
et la surveillance du contenu de la boite à nid avec retrait
éventuel des crottes maternelles, elle a bien confirmé
l'émission puis la disparition de ces crottes laissées
par la mère. La flore digestive des lapereaux a été
analysée tous les 2 jours entre 2 et 10 jours d'âge
par la détermination du nombre de bactéries des genres
Bacteroïdes, Lactobacillus, Streptococcus et Escherichia
coli. Chez les lapereaux en allaitement libre la colonisation
du cæcum par le genre Bacteroïdes (et la quasi
disparition concomitante des genres Lactobacillus et E.
coli) est un peu plus rapide que chez les lapereaux en allaitement
contrôlé une fois par 24h. Le retrait scrupuleux des
crottes laissées par la mère ne fait que retarder
un peu plus la mise en place de la flore cæcale. Mais à
10 jours les 3 groupes expérimentaux avaient des flores tout
à fait comparables. Au plan pratique cela veut dire qu'un
allaitement contrôlé (tétée 1 fois par
24h) ne modifie pratiquement pas la mise en place de la flore digestive
des lapereaux et peut donc être mis en œuvre si d'autres
critères le justifient.
|
| La
présence d'antibiotiques dans l'alimentation réduit
l'importance de la flore cæcale cultivable, mais certains groupes
de bactéries peuvent ne pas être affectés. |
Deux autres
communications l'une portugaise et l'autre hongroise se sont intéressées
aux conséquences sur la flore de la présence d'antibiotiques
dans l'alimentation : bacitracine [27]
et tiamuline +OTC+diclazuril [7]. Dans les deux cas la présence
d'antibiotiques réduit l'importance de la flore totale cultivable
et celle des colibacilles en particulier. La bacitracine fait pratiquement
disparaître les entérocoques [27].
Par contre, dans l'étude hongroise [7],
le complexe antibiotique utilisé n'a pas modifié le
nombre de Bacteroïdes (non déterminé dans l'autre
étude) . Dans cette même étude [7]
l'addition d'un facteur de croissance non antibiotique (benzoquinones
obtenues par culture de levures sur germe de blé) ne modifie
ni l'ampleur de la flore totale, ni le nombre de Bacteroïdes.
|
| Selon
leur nature, des antibiotiques ajoutés dans l'alimentation
peuvent stimuler ou freiner les fermentations cæcales. Les probiotiques
donnent des résultats intermédiaires. |
La
flore digestive analysée à travers le résultat
des fermentations
Comme cela est
classique depuis plusieurs dizaines d'années, certaines communications
ont présenté des analyses des concentrations dans
le contenu cæcal des produits de l'activité de la flore:
acides gras volatils (AGV), ammoniac, pH.
Ainsi la présence
de bacitracine dans l'alimentation (0,1 kg/t) qui réduit
la flore totale [27], tend par
contre à augmenter la concentration du contenu cæcal
[26] en AGV (80 vs 52 mM/L),
et en particulier celle de l'acide butyrique (10,7 vs 4,6 mM/L)
et à réduire le pH cæcal. L'addition le mannane-oligosaccharides
en lieu et place de la bacitracine conduit à des résultats
similaires quand à composition du contenu cæcal.
A l'inverse le complexe tiamuline+OTC+diclazuril placé dans
l'aliment [7] réduit de
manière drastique la concentration en AGV totaux (43 vs 104
mM/L), en réduisant particulièrement les concentrations
en acétate et en butyrate. Le remplacement du complexe antibiotique
par un activateur de croissance (benzoquinones [voir plus haut])
conduit à un résultat intermédiaire.
En marge de
l'activité directe de la flore, un travail roumain de physiologie
aiguë [3] a montré
qu'un accroissement de la concentration d'acétate, de propionate
ou surtout de butyrate dans le sang périphérique stimule
la sécrétion d'amylase pancréatique. Ainsi
une absorption digestive accrue d'AGV issus des fermentations cæcales
et l'augmentation corrélée des taux sanguins, stimule
l'efficacité de la digestion de l'amidon (meilleure hydrolyse
et production de glucose directement absorbé dans l'intestin
grêle) , privant ainsi les bactéries cæcales
d'une partie du substrat leur permettant de produire ces AGV. Cela
correspond à une système classique de régulation
digestive avec modification du fonctionnement physiologique en amont
du point "critique" (ici le cæcum).
|
|
Mesure
quantitative des nutriments élaborés par la flore
digestive
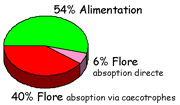
Figure 5
: Origine de la lysine absorbée chaque jour par un lapin
|
La lysine
est un acide aminés indispensable que le lapin doit trouver
que dans son alimentation ou dans les produits synthétisés
par sa flore digestive. Mais en aucun cas la lysine ne peut être
"fabriquée" par le lapin lui-même. En incorporant
du chlorure d'ammonium marqué à l'azote 15 (15N)
dans l'alimentation et en récupérant la lysine totale
et celle marquée 15N dans
les corps bactériens (contenu digestif et cæcotrophes)
mais aussi dans l'organisme du lapin (muscles, foie) il est possible
de déterminer la contribution de la synthèse microbienne
à l'approvisionnement en lysine du lapin. Avec cette technique,
une équipe espagnole de Saragosse [13]
a pu déterminer d'abord que la flore cæcale du lapin
synthétise effectivement de la 15N-lysine
(forme organique) à partir de l'azote 15 du chlorure d'ammonium
(forme minérale). Au plan quantitatif, la lysine synthétisée
par la flore digestive représente 46% de la quantité
de lysine absorbée chaque jour par un lapin. Sur ce total
40% sont absorbés après ingestion des cæcotrophes,
tandis que 6% sont absorbés directement sans passer par l'ingestion
des cæcotrophes. La teneur en 15N-lysine
de la flore cæcale est d'ailleurs indépendante du fait
que les cæcotrophes soient absorbés ou non. Au plan
pratique, cela veut dire que chez tout lapin au fonctionnement digestif
perturbé entraînant un arrêt de l'ingestion des
cæcotrophes (cas très fréquent), il manque 40%
de l'apport de lysine. Cet acide aminé étant indispensable,
le lapin se trouve donc immédiatement carencé (pas
de stockage) et toutes les synthèses protéiques sont
altérées : aussi bien celles nécessaires à
la croissance musculaire ou fœtale que la synthèse des
enzymes nécessaires à l'organisme ou que la synthèse
des immunoglobulines. Ce raisonnement est applicable aux autres
acides aminés indispensables tels que la acides aminés
soufrés ou la thréonine, mais l'ampleur de l'apport
via la flore n'a pas été chiffré.
|
| La
flore digestive synthétise environ 12% des lipides ingérés
chaque jour par un lapin. Elle fournit en particulier jusqu'à
30 mg de CLA par jour (acides linoléniques conjugués) |
Dans
un esprit similaire, une autre équipe espagnole, de Madrid
cette fois [20], a mesuré
la quantité de cæcotrophes produite par les lapins en
fonction de la teneur en fibres solubles de l'alimentation (7,9 à
13,1%, en substitution à de l'amidon - teneur constante en
NDF). La quantité produite s'accroît avec la teneur en
fibres solubles, de 13,6 à 17,7 g de matière sèche
par jour. Parallèlement la quantité de lipides ingérés
en provenance des cæcotrophes s'accroît aussi, passant
de 0,35 à 0,44 g/jour. La composition de ces lipides est également
affectée par les fibres solubles, substrat privilégié
de la flore cæcale. Ainsi la proportion d'acides gras à
nombre impair de carbones (issus exclusivement de la synthèse
bactérienne) passe de 7,2% à 11,4% des acides gras totaux.
De même, les acides gras ramifiés, également issus
de la synthèse bactérienne, passent de 6,7% à
9,7% des acides gras totaux. Enfin les acides gras conjugués
(a. linoléiques conjugués = CLA) augmentent aussi de
6,7 à 8,2% des acides gras totaux. En moyenne, les auteurs
de cette communication [20] estiment
que 12% des lipides ingérés par un lapin proviennent
des synthèses effectuées par sa flore digestive. L'ingestion
de CLA (impliqués dans différents mécanismes
de défense) peut représenter jusqu'à 30 mg par
animal et par jour avec le régime le plus riche en fibres solubles.
Là encore au plan pratique on ne peut que souligner le déséquilibre
nutritionnel engendré par un arrêt de la cæcotrophie
immanquablement associé à toute perturbation du fonctionnement
digestif. |
| |
|
| |
Suite
de l'analyse des communications sur la physiologie digestive

|
| |
|
|
|