| |
|
Sources
de variation de la digestibilité des aliments

|
|
Méthodologie
de détermination du CUDa
La grande majorité
des expérimentateurs détermine la digestibilité
globale des aliments par mesure directe des quantités ingérées
et excrétées (collecte totale des crottes) selon une
méthodologie qui a été parfaitement bien définie,
dite méthode européenne standardisée (Perez
et al., 1995). Toutefois une équipe brésilienne
[25] a cherché à
déterminer la digestibilité des aliments par utilisation
d'un marqueur interne ajouté, en l'occurrence de la lignine
purifiée. Cette méthodologie présente l'avantage
de ne pas nécessiter la mesure de l'ingestion, ni de collecter
toutes les crottes émises. Par contre, elle suppose que la
composition des crottes est constante sur la journée, que
le transit du marqueur est identique à celui de l'aliment,
qu'il n'est ni absorbé ni modifié lors du passage
dans le tube digestif et enfin elle nécessite de doser avec
grande précision le marqueur utilisé dans l'aliment
et dans les crottes. Dans l'expérience présentée
dans cette communication [25],
la mesure de la digestibilité de 5 aliments a été
effectuée en double, par collecte totale et en utilisant
la lignine comme marqueur interne. Pour 2 des 5 aliments, les résultats
ont été comparables, mais pour les 3 autres une différence
significative a été obtenue entre les 2 méthodes
(sous-estimation avec le marqueur interne). La détermination
de la digestibilité en utilisant de la lignine purifiée
comme marqueur interne ne peut donc être retenue.
Dans leur travail
sur la détermination de la valeur alimentaire d'une légumineuse
arbustive des déserts froids (Caragana
microphylla) l'équipe chinoise de Ren et al. [9]
a déterminé la digestibilité des aliments expérimentaux
en utilisant l'insoluble chlorhydrique (minéraux insolubles
dans l'acide chlorhydrique concentré 4M) comme marqueur interne,
sans comparer ses résultats à ceux obtenus avec une
collecte totale. La forte variation de la digestibilité mesurée
en fonction du taux d'incorporation du produit testé (0 -
10 - 20%) et en fonction du type de lapin (jeunes ou adultes) laisse
penser que, comme cela avait été démontré
il y a fort longtemps pour différentes espèces animales,
l'usage de l'insoluble chlorhydrique pour estimer la digestibilité
des aliments est une méthode insuffisamment fiable pour être
recommandée.
|
La
digestibilité apparente fécale des aliments solides
diminue chez les lapereaux entre les âges de 18-20 jours
et celui de 32-35 jours, que les lapereaux soient ou non sevrés.
Elle se stabilise ensuite pour un certain temps, puis tend à
réaugmenter légèrement.
|
Effets
de l'âge ou du type de lapin sur la digestibilité des
aliments
Comme cela l'avait
été par l'équipe françaises de l'INRA-Toulouse
il y a déjà quelques années (Debray
et al., 2000) pour des lapereaux sevrés à 32 jours
et étudiés entre 18 et 42 jours, l'équipe de
Madrid [22] a montré que,
chez les lapereaux sevrés à 25 jours, la digestibilité
de la matière sèche décroît de 25 à
32 jours puis se stabilise entre 32 et 40 jours (CUDa = 69,4 ±
0,47%). Selon ces auteurs, la décroissance entre 25 et 32
jours serait linéaire (- 2,17 points par jour). Le travail
toulousain avait d'ailleurs montré que cette décroissance
concernait l'ensemble des nutriments à l'exception de l'amidon,
quelque soit le ratio amidon/fibres.
Un travail chinois
proposé par Li et al. [23]
a montré que la digestibilité des différents
nutriments est plus élevée (+2 à +4 points)
chez des lapins âgés de 2-3 mois que chez ceux de 1
à 2 mois, l'âge n'étant pas fourni avec plus
de précision. Un résultat similaire a été
observé par une équipe tchèque [28]
ayant mesuré la digestibilité au cours de 3 périodes
: 42-49j puis 49-56 j et enfin 56-63 j.
De leur côté
Gidenne et Fortun-Lamothe [19]
ont montré qu'à l'âge de 6 semaines la digestibilité
des différents nutriments est identique chez des lapins sevrés
à 23 jours et chez des lapins sevrés à 32 jours.
Enfin, en complément de la partie consacrée à
l'effet de l'âge, le travail espagnol précédent
[22] a permis de montrer que
si les CUDa peuvent varier entre individus, cela n'est pas relié
à leur portée d'origine, malgré des différences
très importantes d'ingestion entre individus issus de portées
différentes (5 portées testées).
|
|
Tableau 1
: Digestibilité des nutriments d'un aliment supplémenté
ou non par de la Bacitracine-zinc (0,1 g/kg) chez des lapins étudiés
entre 42 et 46 jours, selon Pinheiro et al. [27]
|
CUDa
|
Aliments
|
Signif.
|
|
Témoin
|
Bacitr.
|
|
M.Sèche
|
70,6
|
69,4
|
ns
|
|
M.Orga
|
79,5
|
78,2
|
P=0,07
|
|
NDF
|
45,4
|
46,0
|
ns
|
|
Azote
|
75,2
|
75,7
|
ns
|
|
Amidon
|
98,3
|
98,3
|
ns
|
|
Lipides
|
88,4
|
87,5
|
ns
|
|
Effets
de l'alimentation sur l'efficacité de la digestion
Comme cela avait
été déjà largement prouvé, l'équipe
tchèque déjà mentionné
[28] a montré qu'une restriction alimentaire quantitative
permet d'améliorer significativement la digestibilité
des aliments, mais cet effet cesse dès que les lapins sont
réalimentés à volonté.
La présence
dans l'alimentation de bacitracine qui, nous l'avons vu plus haut
réduit l'importance de la flore [27]
ne modifie pas la digestibilité des différents nutriments,
en particulier celle des fibres. Seule la digestibilité de
la matière organique est légèrement réduite,
à la limite du seuil de signification (tableau 1 ci-contre).
Selon un travail
présenté par une équipe égyptienne [11],
l'addition d'extraits de yucca (250 mg/kg) ou de lactobacilles vivants
(0,5mg/kg de Lact-A-Bac) permettrait d'améliorer la digestibilité
des différents éléments nutritifs. La conséquence
a été une amélioration de 2 à 5% de
la valeur énergétique de l'aliment de base. Ces additions
permettraient également d'améliorer la vitesse de
croissance (31,6 et 30,9 g/jour vs 28,2 pour le témoin)
ainsi que l'indice de consommation (3,61 et 3,60 vs 3,87).
|
Un accroissement
du taux de fibres (NDF) dans l'alimentation réduit la digestibilitéde
la matière sèche et des protéines, mais sans
modification de celle des fibres elle-même.
|
De manière
classique Nicodemus et al. [24]
ont montré qu'une réduction de l'apport de fibres
alimentaire (25% vs 30% de NDF) se traduit par une meilleure
digestibilité de la matière sèche et des protéines
tant au niveau iléal que fécal. Par contre, la digestibilité
fécale des fibres (ADF 22,0% et NDF 32,9%) n'est pas affectée
par leur taux dans l'aliment. Il faut cependant signaler que cette
diminution du taux de fibres se traduit aussi par un accroissement
significatif de la mortalité des lapins suivis en engraissement
entre 25 et 55 jours (17% vs 8%) sans modification de la
digestibilité iléale ou fécale de l'amidon
(96,6 et 100% respectivement). Rappelons que le taux de fibre le
plus faible se traduit aussi par une forte augmentation de la biodiversité
au sein de la flore cæcale (+ 45% [24]).
Dans ce même travail les auteurs ont montré qu'un broyage
grossier de l'aliment (grille de 9 mm vs grille de 1 mm)
ne modifie pas de manière significative la digestibilité
de la matière sèche ou des protéines, mais
réduit celle des fibres : -8,5% pour NDF et -16,5% pour ADF.
Rappelons que les travaux antérieurs conduits en France sur
les effets du broyage des matières premières entrant
dans les aliments n'avaient pas permis de démontrer de variation
de la digestibilité des aliments, sauf dans le cas d'une
broyage très fin utilisant une grille expérimentale
ayant des orifices de 0,25 mm de diamètre.
|
|
Fonctionnement
du système digestif en relation avec l'âge, le sevrage
et la composition de l'alimentation
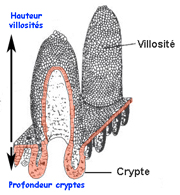
Figure 6
: Vue schématique de la muqueuse intestinale montrant les
villosités et les cryptes
|
Le travail présenté
par Gallois et al. [4]
a permis de montrer que l'âge au sevrage (21 ou 35 jours)
n'a pas d'incidence sur le développement de la muqueuse intestinale
mesuré chaque semaine entre 14 et 49 jours. Par contre la
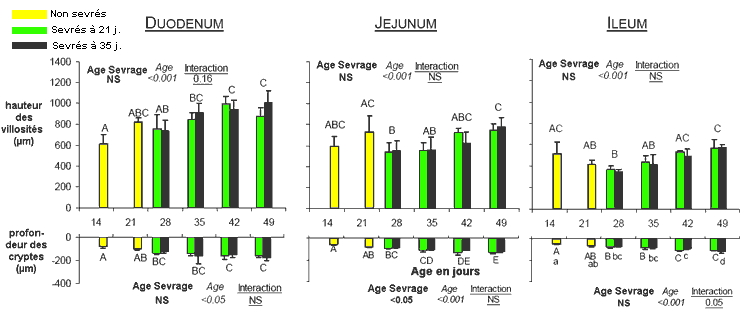
hauteur des villosités intestinales (figure 6 ci-contre)
en 3 points au long de l'intestin grêle (duodénum,
jéjunum et iléon) varie très significativement
avec l'âge (figure
7). Ainsi la hauteur des villosités s'accroît assez
régulièrement avec l'âge dans le duodénum
(de 600 µm à 14 jours jusqu'à 1000 µm
à 42-49 j). Elle diminue à l'inverse dans l'iléon
entre 14 j. (500 µm) et 28 jours (400 µm) pour augmenter
ensuite jusqu'à 49 jours (650 µm), sans relation avec
l'âge des lapins au moment de leur sevrage. La profondeur
des cryptes situées entre les villosités varie aussi
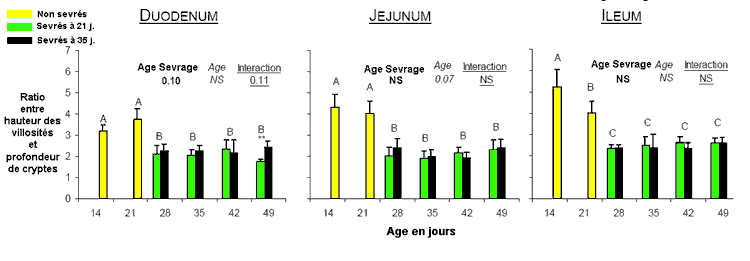
significativement en fonction de l'âge des lapereaux. Tout
au long de l'intestin grêle la hauteur des villosités
représente environ 3 à 5 fois la profondeur des cryptes
à 14 et 21 jours (période d'allaitement strict) alors
qu'elle représente 2 à 2,5 fois la hauteur des cryptes
à partir de 28 jours, sans évolution significative
ni avec l'âge, ni en fonction de l'âge au sevrage (figure
8).
|
|
|
|
Figure
7 : Evolution entre 14 et 49 jours de la hauteur des villosités
et de la profondeur des cryptes en 3 points de l'intestin
grêle des lapins sevrés soit à 21 soit
à 35 jours
|
Figure
8 : Evolution chez le mêmes lapins du rapport
entre la hauteur des villosités et la profondeur
des cryptes.
d'après Gallois et al. [4]
|
|
Tableau
2 : Hauteur des villosités intestinales et digestibilité
iléale de l'amidon en fonction du taux de fibres SOLUBLES
de l'alimentation, chez des lapins sevrés à 25 jours
et sacrifiés à 35.
|
%
Fibres solubles
aliment
|
Hauteur
villosités intestinales
|
Digestibilité
iléale de l'amidon
|
|
7,9
%
|
492
µm
|
93,3%
|
|
13,1
%
|
722
µm
|
93,9%
|
|
Chez des lapins
sevrés à 25 jours et sacrifiés à 35
après avoir consommé un aliment expérimental
contenant 7,9%, 10,3% ou 13,1% de fibres solubles (et 33% de NDF,
20% de protéines et 20% d'amidon) l'accroissement de l'apport
de alimentaire de fibres solubles [21]
entraîne un accroissement de la hauteur des villosités
intestinales(tableau 2). Cet accroissement est associé à
une meilleure digestibilité iléale de l'amidon mais
sans variation significative de la digestibilité iléale
de la matière sèche des aliments.
Le remplacement
d'une partie de l'amidon de l'alimentation des lapereaux sevrés
à 21 jours par de la pectine ou un mélange pectine+inuline
[29], entraîne une plus grande viscosité du contenu
intestinal (P=0,10). De son côté l'apport d'inuline
en plus des pectines semble favoriser les fermentations cæcales,
puisque la concentration en AGV totaux passe de 60 mM/L avec l'aliment
riche en pectines à 93 mM/L lorsque 4% d'inuline sont ajoutés.
|
| La
distribution avant 28 jours d'un aliment pauvre en amidon aux lapereaux,
modifie peu ou de manière transitoire la composition du contenu
de leur cæcum ou leurs performances croissance ultérieures.
Par contre la mortalité 28-70 jours est significativement réduite. |
De 16 à
28 jours d'âge (sevrage) des lapereaux allaités ont
reçu [15] soit l'aliment
maternel (19% d'amidon) soit un aliment dit de sevrage plus pauvre
en amidon (12%). De 20 à 28 jours les consommations de lait
et d'aliment solide ont été identiques dans les 2
groupes et il en a été de même pour la croissance.
A 28 jours les différences de contenu cæcal (quantité,
composition) ont été faibles à nulles. On peut
cependant noter un pH cæcal un peu plus élevé
avec 19% d'amidon qu'avec 12% (6,26 vs 6,05) associé
avec une proportion d'acétate un peu plus faible et une proportion
de butyrate une peu plus forte au sein des acides gras volatils
(25,4% des AGV contre 22,1%). Après le sevrage tous les lapins
ont reçu l'aliment riche en amidon. Ceux qui en avaient eu
peu avant leur sevrage ont eu transitoirement une croissance et
une consommation plus élevées entre 28 et 35 jours,
mais à 70 jours les performances étaient identiques.
La seule différence sensible a été une moralité
28-70 j plus faible avec chez les lapereaux ayant eu peu d'amidon
dans leur jeune âge (3/100 vs 8/100 lapins par lot).
|
|
L'inclusion
de taux croissants de canne à sucre totale (séchée
et broyée, et contenant donc du saccharose) dans l'alimentation
de lapins en croissance se traduit [2]
par une diminution significative de la teneur en matière
sèche du contenu cæcal, mais sans variation significative
de la teneur en protéines ou en fibres de la matière
sèche de ce contenu (11,8% et 6,7% de la MS pour les protéines
et ADF respectivement). La teneur en matière sèche
des fèces diminue de manière drastique avec l'augmentation
du taux de canne à sucre : de 37% avec l'aliment témoin
jusqu'à seulement 18% avec 45% de canne à sucre. On
peut penser que les lapins consommant ce dernier aliment étaient
en permanence à la limite de la diarrhée.
|
Effets
des additions enzymatiques
Un
apport de phytase commerciale (BASF) a permis d'améliorer
l'efficacité de la digestion du phosphore avec une ration
à base de fanes d'arachide, de maïs et de son de riz.
|
Une
idée assez répandue est que les enzymes produites par
l'organisme des animaux et du lapin en particulier, ou par les microorganismes
de leur flore digestive, pourraient ne pas être assez actives
pour hydrolyser la totalité des aliments et permettre l'absorption
maximale des nutriments qu'ils contiennent. Pour améliorer
l'efficacité de la digestion, la solution proposée est
d'ajouter des enzymes exogènes dans l'alimentation, pour suppléer
ou aider celles qui agissent déjà dans le tube digestif.
Avec une ration
à base essentiellement de fanes d'arachide (24%), de maïs
(27%) et de son de riz (18%), et contenant 0,35% de phosphore total,
l'équipe chinoise de Zao et al. [10]
a montré qu'une addition à l'aliment de phytase microbienne
(produite par BASF) permet d'améliorer sensiblement l'absorption
apparente du phosphore total (55% vs 39%) grâce à
une meilleure absorption du phosphore phytique (76% vs 68%).
L'excrétion fécale de calcium, de cuivre, de fer,
de zinc et de manganèse n'ont pas été modifiées.
La vitesse de croissance des lapins rex utilisés pour cet
essai a été améliorée de 11%, sans modification
de l'efficacité alimentaire. Il faut rappeler qu'aucun des
quelques travaux antérieurs conduits sur ce thème
n'avait réellement trouvé d'effet positif lié
à l'addition de phytase exogène, celle produite par
la flore digestive étant trouvée suffisante. Compte
tenu des résultats positifs obtenus par ce travail [10]
il serait intéressant de recommencer l'expérience
avec une ration de base plus proche de celles utilisées en
France ou en Europe de manière générale.
|
|
L'équipe
espagnole de Madrid [18] n'a
trouvé aucune amélioration de performance (vitesse
de croissance moyenne 50 g/j ou indice de consommation moyen de
2,69) après addition soit d'une protéase, soit d'une
protéase + une xylanase dans l'alimentation des lapins en
croissance (origine des enzyme non indiquées dans le texte).
|
|
Les
résultats de cet essai d'addition d'enzymes fibrolytiques
sont trop incohérents pour qu'une conclusion puisse être
tirée
|
Dans un travail
antérieur l'équipe hongroise de Eiben
et al. (2002) avait obtenu des résultats favorables
à la croissance dans les 2 semaines suivant le sevrage, après
addition à la ration des lapins en croissance d'un complexe
de cellulases-hémicellulases (endoglucanases + cellobiohydrolases
+ ß-glucosidases). La même équipe a présenté
lors du Congrès de Puebla une étude [16]
avec des doses réduites du même complexe enzymatique
introduit dans l'alimentation de lapins sevrés à 23
jours (75% - 50% - 25% de la dose antérieure). En remarque
annexe, le poids moyen annoncé par les auteurs pour leurs
lapereaux de 23 jours (510-516 g) nous parait extrêmement
élevé pour des lapins Néo-Zélandais
de cet âge, mais ce sont bien les valeurs écrites dans
la communication. La vitesse de croissance a été
améliorée de manière transitoire, non pas immédiatement
après le sevrage, mais au cours de la période 35-49
jours avec la dose testée la plus élevée (50,7
g/jour avec 53 FPU/kg vs 46,5 g/j pour la témoin -
P=0,067). Cependant, à 77 jours plus aucune différence
n'a été observés entre les différent
lots. De même, l'indice de consommation a été
nettement amélioré au cours de la période 35-49
j (IC 2,59 vs 2,85 - P=0,021) mais détérioré
au cours des 2 semaines suivantes (IC 3,81 vs 3,56 - P = 0,036).
Sur l'ensemble de la période 23-77 jours, seul le lot ayant
reçu l'aliment additionné de 50% de la dose antérieure
a eu un indice de consommation plus faible que le témoin
sans enzymes ajoutées (2,91 vs 3,03 - P = 0,039). Les auteurs
concluent à un effet positif de l'addition du complexe fibrolytique
utilisé en se basant en plus sur la plus faible mortalité
observée après addition d'enzymes à l'alimentation:
6 morts sur 30 lapins en essai dans le lot témoin, contre
4/30 - 4/30 et 1/30 pour les doses croissantes d'enzymes ajoutées.
Malheureusement le test de Khi2
que nous avons fait a posteriori sur ces données montre
que l'écart de mortalité entre le lot témoin
et les 3 lots avec enzymes n'est pas significatif. Nous laissons
donc aux auteurs la responsabilité de l'optimisme de leur
conclusion.
|
Fonctionnement
général de l'organisme et immunité
Les
lapins fabriquent des anticorps circulants spécifiques
de l'aliment qu'ils consomment, mais on en ignore le rôle
physiologique.
|
Un travail préliminaire
(une portée par lot) a été présenté
par une équipe espagnole [14]
sur l'existence d'anticorps sanguins spécifiques à
l'aliment consommé par les lapins. En concevant 2 aliments
de même valeur nutritionnelle mais n'ayant aucune matière
première en commun, distribués de 21 jours à
60 jours aux lapins, les auteurs ont mis en évidence dans
le sang de ces lapins des immunoglobulines de type IgG réagissant
spécifiquement sur des antigènes préparés
avec la partie soluble de chacun des 2 aliments. Les mères
ayant reçu un 3e type d'aliment
ont transmis aux lapereaux (in utero ou via le lait)
des anticorps réagissant aux antigènes issus de ce
3e aliment. La réaction
est très nette chez les lapereaux de 21 jours alors qu'ils
n'ont (en principe) jamais consommé d'aliment solide maternel
ou autre (cage différente de celle de la mère). A
60 jours, la réaction par rapport à l'aliment consommé
en engraissement est toujours forte tandis que celle vis-à-vis
le l'aliment maternel ou de l'autre aliment expérimental
est très atténuée. Pour l'instant on ignore
le rôle joué par ces anticorps, mais on peut supposer
qu'ils ne sont pas élaborés "pour rien".
|
|
Dans le cadre
de l'étude de l'évolution de différents paramètres
digestifs et métaboliques chez des lapins Néo Zélandais
en fonction de l'âge, Li et al. ([23]
- voir plus haut pour l'étude sur la digestibilité)
ont pesé la rate et le thymus, estimant que plus le poids
relatif de ces organes est élevé (en % du poids vif)
plus forte sont les capacités de défense immunitaires
des animaux. Ils n'apportent aucun élément à
l'appui de leur affirmation. Celle-ci mériterait donc une
validation. On ne sait en effet si l'hyper-développement
relatif de ces deux organes est la réponse classique à
une stimulation excessive (début d'infection) ou la preuve
réelle de capacités de réponses immunitaires
supérieures.
|
| Un
complément de sélénium facilement absorbable
a été fourni à des lapins pour stimuler une des
enzymes du métabolisme général. Le résultat
inverse a été obtenu, la complémentation ayant
fait passer l'apport total de sélénium dans la zone
toxique. Une grande prudence est donc recommandée pour l'usage
des additifs alimentaires |
Pour en finir
avec l'analyse des différentes communication présentées
à Puebla en relation avec digestion et le métabolisme
des lapins, il nous semble opportun de rapporter au moins en partie
un travail hongrois [17] dans
lequel les auteurs ont espéré améliorer l'activité
de la glutathion peroxydase (une des enzymes intervenant dans la
limitation de l'oxydation de l'organisme par les radicaux libres
d'oxygène et donc dans celle du processus de vieillissement
et de réaction aux agressions). L'activité de cette
importante enzyme de la famille des peroxydases est "sélénium
dépendante". Les auteurs ont donc espéré
stimuler son activité en ajoutant du sélénium
organique facilement absorbé (Seleno-Yeast®) à
la ration de base : +0,3 mg de Se /kg d'aliment. L'aliment de base
contenait déjà 0,125 mg de Se /kg provenant des matières
premières. La teneur en sélénium a bien augmenté
dans le sang et dans le foie des lapins, mais contrairement à
ce qui était attendu, l'activité de la glutathion
peroxydase a été significativement réduite
dans tous les tissus ou organes où elle a été
mesurée. L'explication est simple: l'apport initial de sélénium
était suffisant et l'apport supplémentaire de sélénium
organique a fait croître l'ingestion total de sélénium
(14 µg/kg vif ) à un niveau proche de celui de la toxicité
(15 µg/kg vif) alors que le besoin reconnu est de 4 µg/kg
vif. Ainsi à vouloir bien faire les auteurs ont dégradé
les défenses des animaux qu'ils espéraient mieux protéger.
Ainsi, même pour les micro-éléments les apports
alimentaires doivent toujours tenir compte de ce qui est déjà
fourni par les matières premières composant les aliments,
et l'usage irraisonné d'additifs alimentaires peut conduire
à l'inverse du résultat escompté.
|
|
|
En conclusion
de cette analyse des communications consacrées à la
digestion et au métabolisme des lapins, côté
négatif on doit remarquer l'absence totale de communication
sur l'estimation in vitro de la digestibilité des
aliments, sujet qui avait fait l'objet de plusieurs interventions
prometteuses lors des précédents congrès.
Côté positif nous devons souligner une fois encore
le foisonnement des idées et des méthodes visant à
mieux suivre la digestion et le métabolisme des lapins. Ces
techniques et méthodes ont été appliquées
quasi exclusivement à des lapins en croissance, mais en commençant
de plus en plus tôt après la naissance.
En particulier ces différents travaux ont permis de démonter
que pour le fonctionnement du tube digestif, l'évolution
de la flore digestive comme pour l'évolution de l'efficacité
de la digestion, le sevrage lui-même (arrêt de l'alimentation
lactée et séparation mère-jeunes) est pratiquement
un non-événement. A l'inverse la connaissance des
différentes évolutions physiologiques peut permettre
de choisir des conditions de sevrage (âge, méthodes)
non plus sur la base de critères subjectifs comme la réduction
d'un stress impossible à quantifier, mais sur la base de
critères objectifs permettant de maximiser les potentialités
ultérieures des lapereaux.
Enfin, l'application
des méthodes de biologie moléculaire à la quantification
de la flore digestive dans différentes situations nutritionnelles
laisse penser que l'augmentation de la biodiversité de la
flore digestive du lapin doit être considérée
comme une dégradation de la situation sanitaire ou tout au
moins comme la preuve d'une aggravation du risque sanitaire.
|
|
|
|
|
{kind=link}